地被菊品种及其近缘种的数量分类1)
2013-08-09孔志新张启翔陈雪娟
孔志新 王 亚 孙 明 张启翔 陈雪娟 刘 华
(北京林业大学,北京,100083)
地被菊是陈俊愉等[1]经过约40 a的育种实践,培育出的一类植株低矮、株型紧凑、观赏价值高、适应性和抗性强、适于地载的新品种群,同时为了便于区别大菊品种群及其盆栽小菊品种群之间的差异,认为凡具有上述特点的菊花品种,可统称之为地被菊[2]。地被菊目前已有百余个品种,在我国绝大部分地区适应生长,尤其在三北地区应用广泛,对低温、干旱、盐碱有较强的抗性。除作为地被植物应用外,其药用、食用价值也不可忽视,一些品种如‘玉人面’‘玉龙’的营养成分和化学成分与‘杭白菊’等传统茶菊基本相似。
地被菊育种的早期亲本主要为野生菊属植物,并随着育种目标的不同加入具有特定性状的亲本。地被菊主要以天然授粉实生选种、人工杂交等为主要育种手段,因此,地被菊品种主要为种间,甚至属间杂交后代,其亲缘关系并不十分明确。数量分类学方法是传统分类学与统计学及计算机技术相结合产生的,可以再现研究对象之间的亲疏关系[3-5],已在慈姑[6]、梅花[7]、杜鹃[8]、芍药[9]、枇杷[10-11]、紫薇[12]等植物资源的品种分类上得到应用。戴思兰等[13]、吴国盛等[14]已采用数量分类方法对部分菊属与亚菊属植物进行聚类分析和亲缘关系的研究,但分类群体的数量有限,并且有关R型聚类分析对各性状间的内在关系还有待进一步深入分析。本研究对46个地被菊品种及其近缘种的表型性状进行聚类分析和主成分分析,旨在分析其亲缘关系,并对各性状之间的内在联系和主要分类指标进行探讨。
1 材料与方法
试验材料取自北京林业大学地被菊资源圃,包括地被菊品种44份,近缘菊属植物2份。将其作为46个分类运算单位(Operational Taxonomic Unit,OUT),详见表1。所有材料均通过温室扦插繁殖,并于5月份移栽至资源圃,在相同环境条件下生长。
分类性状的选取及编码:将所有试验材料进行日常养护管理,根据实地观察测量和常用分类性状,选取31个性状。选取的性状分为3类:数量多态性状17个;定性多态性状12个;二元性状2个。其中29个为形态学性状,2个为生物学性状(自然花期,是否有香味)。性状及其编码见表2。

表1 地被菊品种名及来源
以上性状于2012年8—11月份进行统计,从每个类群中选择5株生长良好、具有代表的植株进行统计,每个性状统计5次,取平均值。
在对数据分析运算之前,先进行标准差标准化处理,以消除不同纲量对数据分析产生的影响[15]。
聚类方法:全部数据采用SPSS18.0软件进行Q型聚类分析,对标准化数据计算各OUT之间的平均欧氏距离[16],然后,采用系统聚类中的离差平方和法[17],做出46个运算分类单位的分类系统树图,最后根据聚类过程中类与类之间的距离测度系数做聚合水平结合线。R型聚类分析方法与之相似,是对OUT的观测量,即分类性状进行分类。
主成分分析:将原来众多具有一定相关性的形态指标,重新组成一组新的相互无关的综合指标,根据每个主成分的方差贡献率来替代原来的指标[18]。并且由因子荷载量得到主成分与对应变量的系数关系。

表2 分类性状及编码
2 结果与分析
2.1 Q型聚类结果分析
46个分类运算单位按欧式平方距离系数由小到大进行聚合,聚合步骤及距离测度系数见表3。从表3可以发现聚类结合过程中最大的跳变出现在第45步,其次是第38步和第42步,由此做3条等级结合线 L1,L2,L3。

表3 Q型聚类进度
由图1可见,等级结合线L1将46个地被菊品种及其近缘种分为两大类群,类群Ⅰ为所有地被菊品种共42份,类群Ⅱ为野生近缘种。类群Ⅰ表现为花色丰富,叶背面多具毛,叶片顶端为深锯齿,植株低矮,舌状花及管状花数量较多且较长,花期长,无香味,观赏价值高。类群Ⅱ包括甘菊(Dendranthema lavandulifolium)与甘野菊(Dendranthemalavandulifoliumvar.seticuspe),其株高均在1 m以上,株幅较大,茎为紫色,叶背面无毛或几无毛,花黄色,舌状花数量小而少,具有香味,花期为10月上旬,观赏价值较低。甘野菊为甘菊的变种,叶片较大。

图1 Q型聚类分析图
结合线L2将Ⅰ类群的42个地被菊品种划分为A、B、C、D 4个类群:A类群包括39个品种;B类群只有1个小菊品种‘香玉’,特点为叶色墨绿,花期晚,为11月上旬,管状花瓣化使其盘花径大于其他地被菊品种,说明小菊与地被菊存在一定差异;C类群由‘密瓣滁龙’‘北极丰’‘淡淡的粉’3个品种组成,它们的共同点是叶长/叶柄长大(平均为5.5/1),其舌状花顶端均为2裂;D类群只有‘阳光小菊’这一品种,其特点十分显著,叶柄较长,达到22.46 mm,因此,叶长/叶柄长较小,叶型狭长,叶色为嫩绿色,其茎节间长、萼片长宽之比均超过其他地被菊品种,管状花数量达到256枚,仅次于‘淡寒粉’。L1将Ⅱ类群全部划分为一类。
等级结合线L3将ⅠA类群进一步划分为a、b、c、d、e、f、g 6 个小组,a有 25 份,b、c有 2 份,d 有 7份,e、f、g各只有1份;将ⅠC类群进一步划分为a、b、c 3 个小组。
ⅠA类群中a组主要为株高0<H≤40 cm,株幅40 cm<D≤80 cm,茎中部粗不超过4 mm的矮生品种。在a组中又结合舌状花颜色、茎色、舌状花顶端形态等进行分类,如‘金毡’‘亮黄早’‘重阳黄’花为黄色,茎为白绿色,舌状花顶端不裂,茎毛量为中等,三者之间亲缘关系较近,欧式距离为14.242;又如‘美矮粉’‘红锦丹’‘夏日’聚在一起 ,表现为叶背面毛多,枝条质地较软,叶长与叶柄长之比接近3.5,舌状花为红或粉红色。b组有2个品种,‘小银球’‘北国之春’,其舌状花数量达到260枚,表现出高度重瓣,管状花少或无,枝条质地较硬。c组包括2个品种,显著的特征为萼片数量多,平均达到95,管状花数量在10以下,舌状花短,仅17.73 mm,茎毛量中等。d组中都是晚花期品种,叶长平均达到48.5 mm,株幅开展,管状花数量较多。e组为‘寒露黄’,虽然其株高,株幅为中等,植株茎粗达5 mm,但叶片大小,花序直径小于其他地被菊品种。f组为‘毛香玉’,与‘寒露黄’相反,表现出花序直径较大,且叶型狭长。g组为‘青莲’,表现为叶片狭长,萼片长度为最小,仅6.28 mm。
ⅠC类群中a组‘密瓣滁龙’与b组‘北极丰’的欧式距离为50.143,与c组‘淡淡的粉’的欧式距离为61.269,可见‘密瓣滁龙’与‘北极丰’的亲缘关系较近。‘淡淡的粉’的花序直径在55 mm以上,舌状花与萼片都超过200枚,而其盘花径却不到5 mm。
从以上聚类结果可以发现,野生菊属植物与地被菊的差异较为明显,主要体现在与观赏性状相关的方面。在地被菊品种内部欧氏距离最大的是‘寒露黄’与小菊品种‘毛香玉’达到117.032。而42个地被菊品种的平均距离为50.635,说明地被菊品种之间的区别也十分突出。
2.2 R型聚类分析
从R型聚类的树系图(图2)可以看出,31个性状被结合线划分为4大类。A类包括花序径、舌状花长、萼片长、茎毛、叶背面毛、舌状花颜色等11个性状,主要是与花序形态、植株被毛有关的性状;B组包括萼片宽、舌状花宽、叶形3个性状;C类包括株幅、株高、盛花期、叶长、叶宽、叶裂、茎色、茎直径等12个性状,主要是与植株大小、叶片形态和植株茎有关;D类包括舌状花形态、舌状花数、萼片数等5个性状。
从性状间的相关性角度分析,可以发现部分性状之间相关程度较高。花序直径与舌状花长相关系数为0.879;叶长与叶宽相关系数为0.799;节间长与叶裂片顶端形态,以及株幅与盛花期之间虽然没有明显的逻辑关系,但相关系数也在0.700以上,说明它们之间存在一定的遗传规律;株高与茎直径的相关系数为0.685;盘花径与管状花数量的相关系数为0.671;叶面颜色、萼片数量、舌状花颜色、茎色与其他性状的相关系数都小于0.400,说明与地被菊各部分颜色的表现较为独立,没有明显的规律。由此可见,多数性状较为独立,且没有明显的负相关性。

图2 R型聚类分析图
2.3 主成分分析
从R型聚类可以看出部分性状之间相关程度较高,为了明确众多分类性状中较为重要的信息,通过几项综合指标实现分类的目的,有必要进行主成分分析。

表4 主成分的特征值、贡献率和累计贡献率
主成分分析结果(表4)表明,主成分贡献率比较分散,累计贡献率增长较慢,直到第10个主成分累计贡献率才达到81.18%,说明地被菊性状变异向多方向发展。第1主成分占信息量的21.33%,前5个主成分占信息总量的59.00%,故选取前5个成分做进一步分析。将因子荷载矩阵与性状抽提系数综合起来[19],得到关于地被菊31个特征形状的总载荷量和性状重要性排名,详见表5。
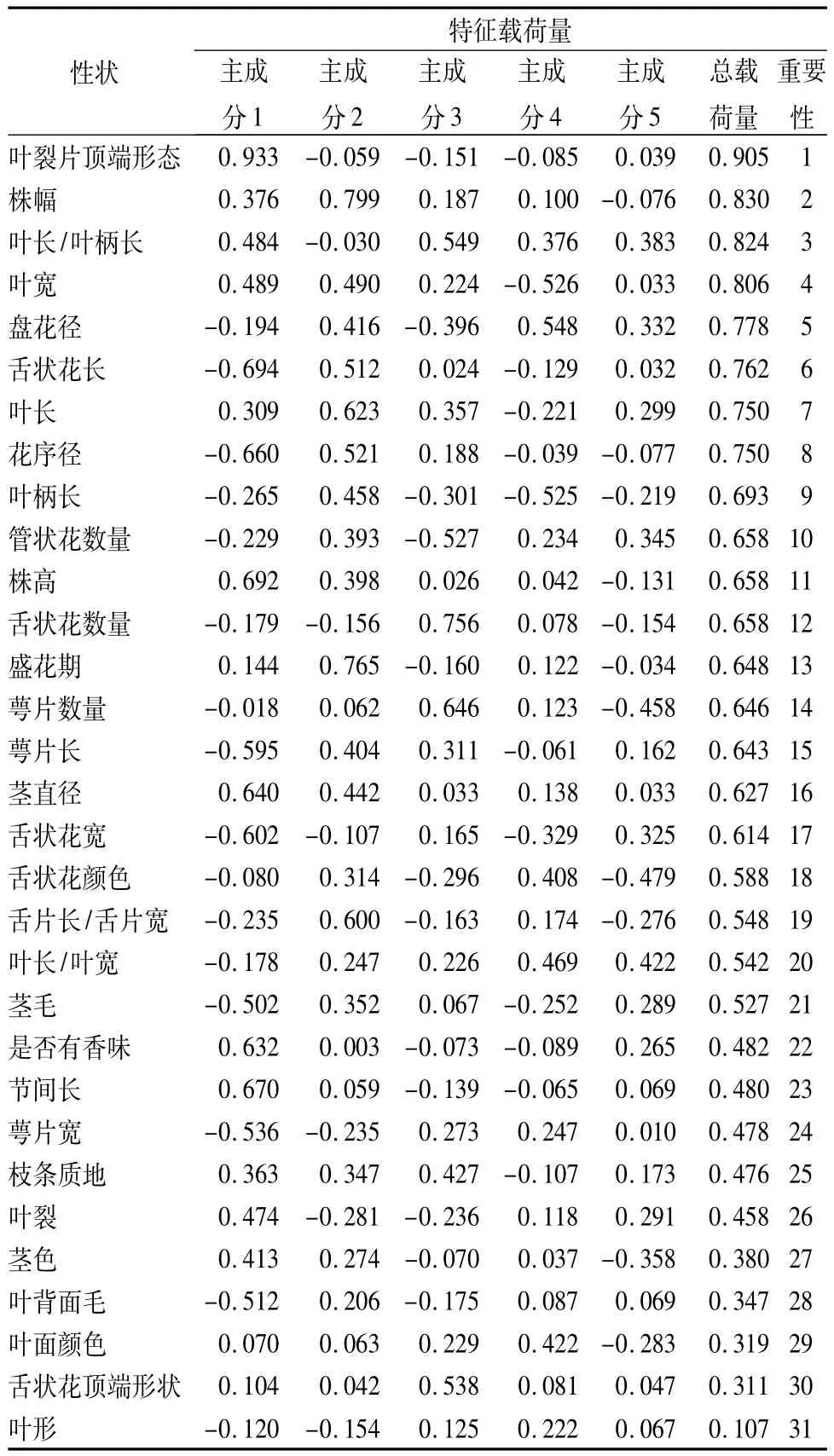
表5 各性状在主成分分析中的载荷量
第1主成分贡献率为21.33%,其中叶裂片顶端形态的载荷量为0.933,舌状花长度、株高、节间长、花序茎、茎直径、是否有香味、舌状花宽7个性状也表现出较大载荷量,绝对值都在0.602以上,这些性状的信息荷载量最大,对分类最为重要。第1主成分主要反映叶裂片顶端为浅锯齿或深锯齿,以及花序、花瓣、植株体量等的尺寸信息。第2主成分贡献率为14.62%,株幅、盛花期、叶长的载荷量较大,均在0.600以上,都是R型聚类中C组的性状,说明这些性状对地被菊分类较为重要。第3主成分贡献率为10.12%,其载荷量绝对值较大的是舌状花数量、萼片数量、管状花数量,叶长/叶柄长、舌状花顶端形状,为代表花序各部分数量的因子。第4主成分的贡献率为6.84%,其载荷量绝对值较大的是盘花径、叶宽、叶长、叶长/叶宽,主要是与叶片尺寸有关的性状。
从性状排序(表5)上看,对地被菊分类最重要的形态性状是叶裂片顶端形态,其次是株幅、叶长/叶柄长、叶宽、盘花径、舌状花长、叶长、花序径,其总荷载量都在0.750以上,重要性占据了所有性状的前8位;重要性排后10位的分别为:是否有香味、节间长、萼片宽、枝条质地、叶裂、茎色、叶背面毛、叶面颜色、舌状花顶端形态、叶形总荷载量都在0.500以下,对分类贡献最小。
3 结论与讨论
地被菊是在岩菊(该名称1963年提出,现不再使用)的育种工作基础上得来的。为克服岩菊株型较高易倒伏的缺点,选取由美国早小菊种子实生苗筛选出的品种“美矮粉”作为母本,与野生种和半野生种,以及紫花野菊、毛华菊等通过人工杂交,在1987年获得第一批地被菊品种。截止2003年,已经培育出6批地被菊品种。在陈俊愉的指导下王彭伟[2]进行了第一二批品种选育天然授粉后代,第一批新品种包括‘铺地雪’‘满天星’‘圆叶白’‘爱黄’等9个。王四清[20]自1988年开始从事第三四批地被菊品种的选育,共培育出23个新品种。本研究中选取的‘金不换’与‘美矮黄’为第三批地被菊品种,且都为‘美矮粉’天然授粉杂交后代,通过Q型聚类发现,‘金不换’与‘美矮黄’被聚为一类,其欧式遗传距离仅为24.914,‘金不换’与‘美矮粉’的遗传距离为34.397。‘旗红’为第4批品种,也为‘美矮粉’天然授粉杂交后代,它们之间的欧式遗传距离仅为35.304,同样,‘金不换’‘美矮黄’‘旗红’三者间的平均遗传距离较近,为31.315。这说明Q型聚类能够对地被菊品种衍化分析起到一定作用,遗传距离较近的品种间可能存在亲缘关系。
孙明[21]曾经利用扩增片段长度多态性(AFLP)技术对39个地被菊品种及小菊品种和18个菊属野生种亲缘关系进行分析,发现地被菊品种两个类群间表现出较远的亲缘关系。在几种聚类方法中主要地被菊品种聚为一类,亲缘关系较近,但是‘美矮黄’‘晚粉’‘红橙早’等 7个地被菊品种,以及‘香玉’‘明星’2个小菊品种与大部分现有地被菊品种有较大的遗传距离,而与大部分野生菊属植物遗传距离较小。本研究选取44个地被菊品种及小菊品种和2个近缘种,通过31个形态性状进行聚类得到了类似的结果。近缘种甘菊、甘野菊虽然是地被菊的主要亲本,但在地被菊长期选择育种过程中有诸多亲本参与其中,使部分地被菊品种与其原始亲本形态间差别逐渐加大,因此,近缘种聚为一类,品种聚为一大类。Q型聚类图中,小菊品种‘香玉’以及地被菊‘密瓣滁龙’‘北极丰’‘淡淡的粉’与多数地被菊品种的亲缘关系较远,而它们与其他地被菊的差别主要表现在盘花直径以及叶长/叶柄长。
传统的菊花分类方法,按照花色、花期、花朵直径、舌瓣瓣形进行分类[22],并不完全适用于地被菊的分类,因为地被菊都是小菊类,舌瓣多数为平瓣类,花期又普遍较大菊早。戴思兰等[23]认为,菊属植物性状间的相关性不强,性状分布较为分散,没有明显的分成几大组。本试验通过R型聚类和主成分分析,将31个性状分为4大类,其中A组和C组中性状具有一定的关联性,而B组和D组中的性状几乎没有相关性。这也说明分类性状的选择并非越多越好,一部分性状,萼片宽、枝条质地、叶裂、茎色、叶背面毛、叶面颜色、舌状花顶端形态等在地被菊分类上作用很小,应该选择重要性状,首先注意叶裂片顶端形态,其次是株幅、叶长/叶柄长、叶宽、盘花径、舌状花等,避免过多次重要性状的干扰。
以上只是从形态学方面对地被菊品种进行初步的分类,要得到更准确的分类结果还需要收集更多的品种,并且连续多年对形态性状进行统计,以消除气候造成的影响,同时还可以与分子标记手段相结合。
[1]陈俊愉,崔娇鹏.地被菊培育与造景[M].北京:中国林业出版社,2006.
[2]王彭伟.地被菊选育的研究[D].北京:北京林业大学,1988.
[3]赵冰,雒新艳,张启翔.蜡梅品种的数量分类研究[J].园艺学报,2007,34(4):947-954.
[4]Stace C A.Plant taxonomy and biosystem atics[M].London:Edw ard Arnold,1980.
[5]Farris J S.Methods for computing wagner trees[J].System atic Zoology,1970,19:83-92.
[6]陈家宽,孙祥钟,玉徽勤,等.中国慈姑属的数量分类研究[J].武汉大学学报:自然科学版,1988(1):107-114.
[7]杨果,李彦,吕英民,等.梅花品种数量分类研究[J].北京林业大学学报,2010,32(2):46-51.
[8]周兰英,王永清,张丽,等.46种杜鹃花属植物表型性状的数量分类[J].林业科学,2009,45(8):67-75.
[9]郭先锋,王莲英.我国栽培芍药与几个近缘种的数量分类研究[J].园艺学报,2005,32(3):473-476.
[10]陈秀萍,黄爱萍,蒋际谋,等.枇杷种质资源数量分类研究[J].园艺学报,2011,38(4):644-656.
[11]Leguizamòn JE,Badenes M L.Multivariate analysis as a tool for germplasm studies,example of analysis of germplasm loquat data[J].ActaHorticulturae,2003,606:29-35.
[12]顾翠花,王守先,田苗.紫薇品种数量分类研究[J].浙江林学院学报,2010,27(6):903-907.
[13]戴思兰,陈俊愉.中国菊属一些种的分支分类学研究[J].武汉植物学研究,1997,15(1):27-34.
[14]吴国盛,陈发棣,陈素梅,等.部分菊属与亚菊属植物的形态学聚类及亲缘关系分析[J].南京农业大学学报,2009,32(1):155-159.
[15]徐克学.生物教学[M].北京:科学出版社,1999.
[16]胡文舜,陈秀萍,李韬,等.云南部分野生枇杷种质资源的数量分类初探[J].果树学报,2009,26(3):403-408.
[17]刘权,王桂荣,吕均良,等.枇杷品种资源的数量分类[J].果树科学,1993,10(3):137-141.
[18]李朝峰,杨中宝.SPSS主成分分析中的特征向量计算问题[J].统计教育,2007,90(3):10-11.
[19]万静.48种悬钩子属植物表型性状数量分类研究[D].雅安:四川农业大学,2010.
[20]王四清.地被菊遗传育种研究[D].北京:北京林业大学,1993.
[21]孙明.地被菊及近缘种亲缘关系分析和新品种选育[D].北京:北京林业大学,2007.
[22]段东泰,高全荣.菊花观赏与培育[M].北京:农村读物出版社,2001.
[23]戴思兰,钟扬,张晓艳.中国菊属植物部分种的数量分类研究[J].北京林业大学学报,1995,17(4):9-14.
