不同环境下黄山杜鹃光合特性及其与主要环境因子的关系1)
2013-08-09孟艳琼程耀辉闫士凤秦金舟
孙 婧 王 刚 孟艳琼 程耀辉 闫士凤 秦金舟
(安徽农业大学,合肥,230036)
黄山杜鹃(Rhododendronanhweiense),即安徽杜鹃,又名皖杜鹃,是杜鹃花科杜鹃花属常绿灌木。1986年3月1日,安徽省人大六届二十次常委会将其定为省花;1987年在江苏无锡举办的首届中国杜鹃花展览会上被评为“最佳原种奖”[1]。黄山杜鹃原产安徽黄山、清凉峰、枯牛降、天柱山等地,浙江、江西等地也有分布,多生长于海拔1 000 m以上的岩坡灌丛中,生长适温为10~25℃[2]。黄山杜鹃花大色艳,其植株不仅具有极强的观赏性,并且能够吸收空气中臭氧、二氧化硫等有害气体,是具有广泛应用前景的盆栽观赏和园林绿化树种。目前对于黄山杜鹃的引种研究报道较少,研究内容仅为引种后实生苗生长状况以及播种出芽情况[3],而目前对于不同栽培环境下黄山杜鹃植株的光合作用等生理因素的影响还未见报道。影响植物的光合作用有许多因子,包括光补偿点、CO2浓度等[4],环境的变化可以引起植物净光合速率以及其他生理过程的变化[5]。本试验通过比较3个不同环境下的黄山杜鹃引种苗光合特性及相关因子的差异,研究环境因子对于黄山杜鹃光合生理特性的影响,从而为黄山杜鹃引种工作提供一定依据。
1 试验地概况
试验地分别位于合肥市半岛苗圃、合肥市大杨镇和安徽农业大学农萃园。合肥地处中纬度地带,全年气侯冬寒夏热,春秋温和,为亚热带湿润季风气候。年平均气温15.7℃,年降水量近1 000 mm,日照时数达2 100 h。其中半岛苗圃距离市中心约13 km,园区西、北、东三面为董铺水库环绕,园区内种植大量乔灌木,园区内小气候较外界温和,黄山杜鹃栽培于水杉林下;大杨镇距离市中心约13.5 km,黄山杜鹃植株周围无遮挡物,栽培地周围无较大水系;农萃园距离市中心2.4 km,校园内树木繁盛,小气候温和。
2 材料与方法
供试苗木为黄山杜鹃,来源于安徽黄山风景区,植株年龄均为5 a以上的成年植株,引种栽培时间为3 a。光合指标测定于2010年4月中旬进行。
于天气晴朗的2010年4月17日至4月20日,采用Li-6400便携式光合作用测定仪,分别测定黄山杜鹃在不同栽培环境下的光合日变化,同时测定当日各环境因子的变化;测定时选取生长健康、生长部位和生长态势相对一致的向阳叶片[6],本试验分别选取黄山杜鹃植株上部受光较好的健壮叶片进行测定,从08:00—16:00每隔1 h测定1次光合指标,每个处理重复3次。测定指标包括:净光合速率(Pn,μmol·m-2·s-1)、蒸腾速率(Tr,mmol·m-2·s-1)、胞间 CO2摩尔分数(Ci,μmol·mol-1)和气孔导度 (Gs,mol· m-2· s-1),并记录环境温度(Ta,℃)、空气相对湿度 (RH,%)、有效光辐射(PA,R,μmol·m-2·s-1)等环境因子指标。
3 结果与分析
3.1 不同栽培环境下环境因子日变化
植物的光合作用受到多种因子的影响,包括植株的内在生理因子和外部的环境因子共同作用。由于本次试验中的黄山杜鹃引种苗树龄相近且移栽年限也相近,因此,对于其内在因子不作考虑,仅比较三处栽培地点的环境因子差异。
从表1可以看出在不同的栽培环境中,大杨镇的有效光辐射(PA,R)和环境温度(Ta)都明显高于半岛苗圃和农萃园,而环境湿度(RH)却明显小于其他两地区。由表2对于环境因子间的相关性分析可以得出RH和Ta为显著负相关(p<0.01),RH与PA,R也呈现显著负相关(p<0.01),而PA,R与Ta则为正相关,且PA,R受RH影响较大。
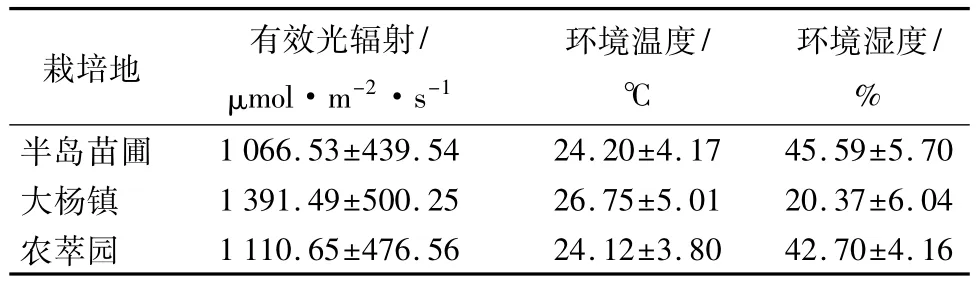
表1 不同栽培环境下环境因子差异
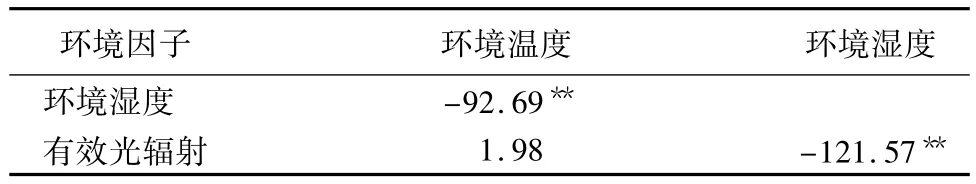
表2 不同栽培环境下环境因子间相关性分析
由图1可知半岛苗圃、大杨镇和农萃园三地区的Ta均从08:00开始逐渐升高,全天最高温度分别为 31.16、34.74、29.56 ℃,之后Ta均呈下降趋势(图1a);RH日变化曲线均为两头高中间低的凹型曲线,在午间到达最低值,分别为36.16%、12.53%、35.16%(图1b);PA,R日变化曲线均为“凸”型曲线,于11:00—13:00 达到全天最高,13:00 以后PA,R开始下降(图1c)。

图1 不同栽培环境下环境因子日变化
3.2 不同栽培环境下黄山杜鹃光合特性日变化
3.2.1 净光合速率日变化
由图2 a可知,不同栽培环境下的黄山杜鹃净光合速率(Pn)均呈现“M”型双峰曲线,并且第一峰值均高于第二峰值,大杨镇和农萃园的Pn日变化幅度较大,而半岛苗圃的日变化幅度较小。大杨镇和农萃园的Pn在10:00达到第一个峰值,分别为 3.450、4.620 μmol·m-2·s-1,随后开始下降,并分别在12:00和13:00达到峰谷,分别为0.453、0.862 μmol·m-2·s-1;半岛苗圃的Pn在 11:00 达到第一峰值,为 1.683 μmol·m-2·s-1,在 13:00 降至峰谷,为 0.345μmol·m-2·s-1。大杨镇和农萃园Pn在15:00 达到第二峰值,分别为 1.383、3.120 μmol·m-2·s-1;半岛苗圃的Pn在14:00达到第二峰值,为0.953 μmol·m-2·s-1。结合图1 可以发现,三处地区午间的Ta和PA,R较高、空气湿度较小,黄山杜鹃在12:00—13:00出现光合“午休现象”与环境因子有紧密联系。从图1和图2a看出,半岛苗圃和农萃园的环境因子虽然较为相近,但其Pn差别较大,造成这种现象的具体原因还需做进一步研究。影响Pn的主要环境因子为PA,R和Ta。
3.2.2 蒸腾速率日变化
由图2b可知,3条蒸腾速率(Tr)日变化曲线均为“凸”型,其中大杨镇全天Tr明显要高于其他两地;半岛苗圃Tr日变化最为平缓。3条曲线在08:00—10:00均为缓慢上升状态,大杨镇的Tr在10:00—11:00 蒸腾速率急速上升,达到1.810 mmol·m-2·s-1,并一直保持较高的水平直到14:00,随后急速下降至 0.558 mmol·m-2·s-1,并在 15:00 以后呈现缓慢下降趋势。半岛苗圃的Tr在11:00时达到全天最高点,为 0.516 mmol·m-2·s-1,在 12:00—16:00,其Tr一直处于平稳下降状态;农萃园的Tr峰点比半岛苗圃峰点出现的时间晚1 h,在12:00时达到 0.876 mmol·m-2·s-1,并在之后一直保持下降趋势。
3.2.3 气孔导度日变化
由图2c可知,3条气孔导度(Gs)日变化曲线均呈不规则型,且3条曲线上午的起伏程度均明显小于下午的起伏程度。大杨镇和农萃园的Gs曲线自08:00起呈现下降状态,两地08:00的Gs值分别为0.0434 、0.036 1 mol·m-2·s-1,两地的Gs曲线分别于15:00 和14:00 达到全天最高值,分别为 0.069 8、0.068 1 mol·m-2·s-1;而半岛苗圃的Gs曲线在08:00—13:00 为上升状态,峰值为 0.067 8 mol·m-2·s-1,14:00 降至 0.031 9 mol·m-2·s-1,14:00—16:00Gs曲线均呈现上升状态。结合图2a可以发现,Gs日变化曲线的起伏滞后于Pn日变化曲线的起伏,且08:00—10:00大杨镇和农萃园的Gs曲线与Pn曲线的状态相反,造成这一现象的具体原因还需要进一步研究;午间的光合“午休”现象时,Gs呈现平缓状态,“午休”过后其Gs值增大,气孔张大,Pn也开始增加。
3.2.4 胞间CO2摩尔分数日变化
由图2d可知,半岛苗圃和农萃园的胞间CO2摩尔分数(Ci)日变化曲线均呈两头高中间低的“凹”型曲线,在08:00—11:00,Ci均呈现下降趋势,并在 12:00 降至最低,分别为 146、172 μmol·mol-1;大杨镇Ci在 11:00—12:00略有上升,升至296 μmol·mol-1,随后开始下降。在 14:00—16:00,3条曲线均呈现上升趋势,这可能是由于光照减弱、Gs下降,从而导致细胞内CO2摩尔分数的积累。一般认为影响植物光合作用的因素为气孔因素和非气孔因素,而本次试验结果显示,Gs和Ci变化基本相反,则说明Pn的下降是由于非气孔因素造成的,是由于叶肉细胞光合能力的降低引起的[7]。

图2 不同栽培环境下光合特性日变化
3.3 不同栽培环境下光合特性与环境因子相关性分析
植物Pn是体现光合作用强弱的重要指标,由表3可知,不同栽培环境下Pn与环境因子的相关性不同,半岛苗圃Pn与PA,R的相关性最大,为显著正相关;大杨镇Pn则与Ta为显著负相关,与RH和PA,R为显著正相关;农萃园Pn与Ta为显著负相关,与PA,R为显著正相关。影响黄山杜鹃Pn的主要因子有Ta、RH、PA,R以及Ci。半岛苗圃Pn的影响因子相关性由大到小的顺序为:Ci、PA,R、Tr、Gs、RH、Ta;大杨镇Pn的影响因子相关性由大到小的顺序为:PA,R、RH、Ta、Tr、Ci、Gs;农萃园Pn的影响因子相关性由大到小的顺序为:PA,R、Ta、Tr、Ci、Gs、RH。
4 结论与讨论
本试验中不同栽培地区的黄山杜鹃Pn曲线均为双峰型,在午间气温、光照最强时有“午休现象”,且第一峰的高度明显高于第二峰,说明Pn在午间的下降幅度要明显大于午后的回升幅度。黄山杜鹃午间Pn降低时,Gs和Ci并不是同时呈现下降状态,说明光合速率是受非气孔限制影响。午间Ta和PA,R的增加抑制了黄山杜鹃的Pn,其原因可能是因为高温增加了叶片的光呼吸强度,并导致呼吸消耗的增多,从而造成Pn下降[8]。大杨镇的黄山杜鹃净光合速率要明显小于农萃园,造成此现象的主要原因可能是由于高温增加了叶片的光呼吸强度[9],使得呼吸消耗增多,从而导致净光合速率的降低。

表3 环境因子和光合特性相关性分析
从半岛苗圃和农萃园所测得数据可以看出,虽然其所处环境因子较为近似,但其Pn差别较大,造成这种现象的具体原因还需做进一步研究。影响Pn的主要环境因子为PA,R。
从植株生长状况来看,半岛苗圃和农萃园的黄山杜鹃生长状况较好,叶色均为深绿并有光泽,叶边缘微反卷,芽色多为红棕色,少量为绿色。而大杨镇的黄山杜鹃植株生长状况不佳,叶色偏黄,叶缘严重反卷,芽色多为黄绿色。这可能是由于大杨镇栽培地周围无遮挡物,植株在白天处于较强光照下,并且栽培地周围没有较大水系,所以其整个植株水分散失较大,造成叶片萎蔫下垂。因此,在养护管理方面应采取遮阳措施,避免阳光长时间直射造成水分散失过多。
将黄山杜鹃大面积由高山引种到城市,由于城市环境条件较其原产地炎热、干燥,黄山杜鹃的生长状况很可能会受到影响,因此,应尽量将其栽植在有散射光处,避免其直接处于强光照、高温、低湿的环境中[10-11]。
[1]胡一民,骆绪美,姚剑飞.安徽省花:黄山杜鹃[J].中国花卉盆景,2005(3):4-5.
[2]傅玉兰.黄山杜鹃花科种质资源初探[J].园艺学报,1990,17(1):34-38.
[3]韩九同,陶涛,江朝宝,等.安徽杜鹃的引种繁殖技术[J].经济林研究,2008,26(4):76-79.
[4]Xin Guangzhu,Stephen P L,Donald R O.Improving photosynthetic efficiency for greater yield[J].Annual Review of Plant Biology,2010(61):235-261.
[5]靳甜甜,傅伯杰,刘国华,等.不同坡位沙棘光合日变化及其主要环境因子[J].生态学报,2011,31(7):1783-1793.
[6]张志平,傅玉兰,吴根松,等.天目杜鹃生理生态研究[J].中国农学通报,2006,22(12):99-102.
[7]柯世省.云锦杜鹃夏季气体交换和水分利用效率日变化[J].生物学杂志,2007,24(4):37-40.
[8]Takahashi S,Bauwe H,Badger M.Impairment of thepho torespiratory pathway accelerates photosystemⅡ.by suppression of repair but notacceleration of damage processes inArabidopsis[J].Plant Physiology,2007,144(1):487-494.
[9]Duarte H M,Luttge U.Correlation between photorespiration,CO2-assimilation and spatioternporal dynamics of photosynthesis/crassulacean acid metabolism-intermediate speciesClusiaminorL.(Clusiaceae)[J].Trees,2007,21(5):531-540.
[10]康美丽,李永红,谢丽娟,等.毛棉杜鹃的生物学特性与栽培管理[J].安徽农业科学,2009,37(16):7389-7391.
[11]刁松峰,万志兵,黄欣,等.黄山杜鹃适宜的发芽条件和苗期生物学特性探究[J].林业实用技术,2010(7):49-50.
