攀西地区黑山羊mtDNA D-loop区遗传多态性研究
2013-08-08姜雪鸥韦雷飞钟金城
姜雪鸥,杨 鹰,万 洁,韦雷飞,钟金城,李 娜
(1.西南民族大学动物遗传育种学国家民委-教育部重点实验室,成都 610041;2.攀枝花市农林科学研究院,四川 攀枝花 617061)
黑山羊主要分布在海拔2 500米以下地区,这里年平均气温约为15.2℃,年平均相对湿度为69%~70%,黑山羊体格中等,体躯匀称,略呈长方形。头呈三角形,鼻梁平直,两耳向前倾立,公、母羊绝大多数有角、有髯,公羊角粗大,呈镰刀状,略向后外侧扭转,母羊角较小,多向后上方弯曲,向外侧扭转。毛被光泽好,全为黑色。毛被内层生长有短而稀的绒毛。随着外来品种不断引进以及杂交改良、人工授精和胚胎移植等工作展开,地方品种群体数量急剧下降,遗传多样性降低。
线粒体 DNA(Mitochondrial DNA,mtDNA)是唯一核外遗传物质,由于线粒体高拷贝数、缺乏重组以及单亲及母系遗传等特点而使其成为系统发育和遗传结构研究良好素材。而其中片段长为1 220 bp mtDNA D-环控制区(D-Loop)是mtDNA基因组中进化速率最高、最具多态区域,尤其在高变区,由于D环受自然选择影响较大,存在巨大变异,所以一般用于种群间系统进化分析[1]。
本研究选择攀枝花各区县本地黑山羊,建昌黑山羊,以及周边黑山羊品种(群体)为研究对象,来分析黑山羊在种内遗传结构,探讨攀西黑山羊起源、系统进化关系及其分类地位。
1 材料与方法
1.1 试验动物及基因组DNA提取
随机选取健康成年黑山羊共185个血样,黑山羊品种及数量见表1。使用全基因组DNA提取试剂盒(天根生物公司),提取基因组DNA。

表1 样本信息Table 1 Information of samples
1.2 PCR扩增、克隆及测序
参照NCBI http://www.ncbi.nlm.nih.gov上已有成都麻羊(GenBank Accession No:AF533441)、绵羊(GenBank Accession No:AF039578)、猪(GenBank Accession No:DQ534707)和牦牛(GenBank Acces⁃sion No:NC_006380)mtDNA D-loop区序列,应用生物信息学软件DNAMAN设计引物,由英潍捷基(上海)贸易有限公司合成。引物序列为:PF:5'CATCATTCTAGTAATAATA CC 3'与PR:5'CTTAC⁃CATGTAAAAGACCCAG 3'。
PCR扩增体系:总体系为25 μL,包括10×Ex Taq Buffer(Mg2+Free)2.5 μL, MgCl2(25 mmol·L-1)1.5 μL,dNTPMix-ture(各 2.5 mmol·L-1)2 μL,模板DNA(50 ng·μL-1)2 μL,上、下游引物各1 μL,Ex Taq(5 U·μL-1)0.15 μL,灭菌去离子水14.85 μL。
PCR扩增程序:94℃预变性3 min;94℃变性30 s,62.5℃退火30s,72℃延伸90 s,30个循环;最后72℃总延伸5 min,4℃保存。PCR产物用1%琼脂糖凝胶电泳,GV染色并用凝胶成像系统检测。
根据检测结果,对185只黑山羊mtDNA DLoop区PCR产物用DNA凝胶回收试剂盒(Axygene生物公司)进行胶回收分离纯化,纯化后目片段与pMDTM19-T Vector于16℃恒温金属浴中接4 h;而后将连接产物转化到大肠杆菌DH5α感受态细胞中培养;再将菌液涂布在含有X-Gal底物、IPTG诱导物和氨苄青霉素(Ampr)固体LB平板上,利用蓝白斑遗传学筛选法筛选;用灭菌牙签挑取白色单一菌落,在含氨卞青霉素(Ampr)LB液体培养基中培养并提取质粒,将构建重组质粒通过PCR法转化子鉴定,以确保克隆成功。提取阳性重组 质粒,将其送往英潍捷基(上海)贸易有限公司测序[2]。
1.3 数据处理
测序结果用DNAMAN、BioEdit、DnaSPv5等生物软件进行序列比对分析;用MEGA生物软件构建不同物种间系统进化树。
2 结果与分析
2.1 mtDNA D-loop扩增结果
根据GenBank中绵羊和成都麻羊等序列设计引物,PCR扩增出黑山羊mtDNA D-loop区片段长度应为1 412 bp,结果见图1。
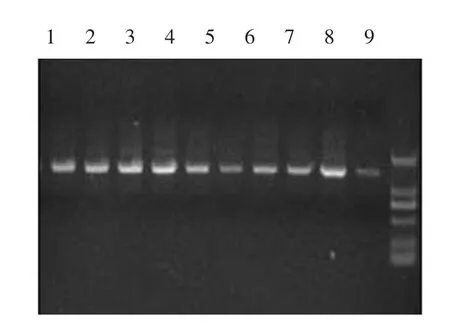
图1 黑山羊mtDNA D-loop区全序列PCR扩增结果Fig.1 PCR amplification of the mtDNA D-loop region of Black goat
其中,M泳道为MarkerD2 000,1~9泳道分别为本地黑山羊(YB、LT、ZB、MY)和周边黑山羊(YL、HL、JC、JT、LZ)PCR产物,本地黑山羊与周边黑山羊mtDNA D-loop区PCR扩增产物大小均在1 412 bp,与预期结果一致。
2.2 mtDNA D-loop区全序列长度与碱基组成
本研究测定黑山羊9个黑山羊类群185个个体mtDNA D-loop区全序列,其长度为1 214~1 224 bp,T、C、A、G 4种核苷酸平均比例分别为28.7%(28.5%~28.89%)、25.9%(25.8%~26.2%)、31.1%(31.0%~31.4%)、14.2%(13.7%~14.5%)。
2.3 mtDNA D-loop区全序列遗传多样性分析
通过DnaSPv5软件分析185头黑山羊,共检测mtDNA D-loop区序列有53种单倍体(见表3),其中HL黑山羊单倍型高于其他8个类群,而MY、YB、ZB黑山羊单倍型数最小,均为3。Tajima's D检验是中性检验一种,LT、LZ、JT、ZBTajima's D统计值为正值或者接近零(P>0.05),说明其Taji⁃ma's D中性检验不显著,序列进化方式为平衡选择,且有一些单倍型分化;而其他7个类群Taji⁃ma's D统计值均为负值(0.01<P<0.05),则说明基Tajima's D中性检验显著,进化方式不平衡,为负向选择,可能由于近一段时期群体扩张产生丰富稀有等位基因[3]。
9个类群mtDNA D-loop区序列多态性结果见表4。

表2 9个类群黑山羊mtDNA D-loop区碱基组成比较Table 2 Base composition of of the mtDNA D-loop region of Black goat

表3 9个类群黑山羊mtDNA D-loop区序列单倍型多态性与核苷酸多样性Table 3 Nucleotide diversity and haplotype diversity of the mtDNA D-loop region of Black goat

表4 9个类群黑山羊mtDNA D-loop区序列多态性Table 4 Polymorphism of the mtDNA D-loop region of Black goat
从表4可知,从群体内分析,LZ、JT、HL、YB、ZB各群体多样性较MY、LT、YL、JC各群体多样性丰富;LZ群体内多态位点最为丰富(117),而MY群体内多态位点最为贫乏(7);LZ群体内简约信息位点最多为65个,其次是JT、HL、LZ、YL、ZB、JC、YB,而MY群体内简约信息位点最少为1个。从群体间分析,所有类群共有178个多态位点,47个单一多态位点,131个简约信息位点。核苷酸歧义度是反映群体内mtDNA多态度一个指标[4-5],在所研究各黑山羊群体中,均出现不同程度变异,群体内mtDNA多态程度高低依次为LZ、JT、HL、YB、ZB、JC、YL、LT、MY。9个类群核苷酸歧义度为0.035 00。
2.4 遗传距离
根据Mega 5软件取得九个群体mtDNA D-loop一致序,并与自GenBank中获取羊族5属mtDNA D-loop区序列(各物种名称、登录号及序列长度见表5),计算遗传距离(D),见表6。

表5 其他物种信息Table 5 Information of other species
从表6可知,各类群之间序列差异在0.000~0.188。其中云岭黑山羊(YL)与大龙潭乡黑山羊(LT)序列差异最小,为0.000,表明亲缘关系最近;其次是攀枝花市建昌黑山羊(JC)分别与云岭黑山羊(YL)、会理黑山羊(HL)、大龙潭乡黑山羊(LT)序列差异都为0.002;而蛮羊Ammotragus ler⁃via与绵羊Ovis aries序列差异最大,为0.188,说明亲缘关系最远。表6也表明,相对于与绵羊Ovis ar⁃ies、野山 羊 Capra aegagrus、蛮羊 Ammotragus ler⁃via、塔尔羊Hemitragus jemlahicus和倭岩羊Pseudo⁃is schaeferi羊族五属关系,9个黑山羊群体之间遗传距离更近,同源性更高。
2.5 聚类结果
分别用邻接法和最小进化法构建9个群体黑山羊与绵羊Ovis aries、野山羊Capra aegagrus、蛮羊Ammotragus lervia、塔尔羊Hemitragus jemlahicus和倭岩羊Pseudois schaeferi羊族五属mtDNA D-loop区序列系统进化树,如图2~3所示,两种方法得出大致相同拓扑结构。结果显示:会理黑山羊(HL)与攀枝花市建昌黑山羊(JC)属于同一个类、大龙潭乡黑山羊(LT)与云岭黑山羊(YL)属于同一类,两个分支先与YB聚在一起为一个分支;此分支和ZB为一合枝;以上聚合枝与野山羊Capra aegagrus聚为一个合枝;JT与LZ聚在一起,与上一合枝聚为1类;1类与MY聚为一大类;依次再与塔尔羊Hemitragus jemlahicus、蛮羊 Ammotragus lervia、倭岩羊Pseudois schaeferi、绵羊Ovis aries聚在一起。9个群体黑山羊是同一属,因而亲缘关系比较近。枝上数值为重复1000次支持率。以上系统进化关系还可以从9个群体黑山羊与羊族五属遗传距离得以验证(见表4)。由图2还可知,9个类群黑山羊明显分为3支系,即支系A~C,支系A包括单倍型数目最多,其次是支系B,支系C只包括1中单倍型。

表6 14个类群mtDNA D-loop区间遗传距离(D)Table 6 Genetic distance among 14 groups of Caprini

图2 用NJ法构建mtDNA D-loop区序列系统进化树Fig.2 NJ phylogenetic tree between 14 groups of Caprini

图3 用ME法构建mtDNA D-loop区序列系统进化树Fig.3 ME phylogenetic tree between 14 groups of Caprini
3 讨论与结论
本试验根据NCBI上已有成都麻羊(GenBank Accession No:AF533441)、绵羊(GenBank Acces⁃sion No:AF039578)、猪(GenBank Accession No:DQ534707)和牦牛(GenBank Accession No:NC_006380)mtDNA D-loop区序列信息设计引物,成功扩增出黑山羊mtDNA D-loop区序列。
该基因长度为1 214~1 224 bp。碱基含量也与其他羊类相近,A+T含量59.8%显著高于G+C含量40.2%。表现出一定碱基偏好性,A+T含量高是基因非编码区普遍特征[2],显示GC→AT突变方向。
衡量一个群体mtDNA遗传变异程度指标,Nei定义是给定群体内两个随机选取mtDNA序列间平均每个位点核苷酸差异数目(歧义度),该值越小,说明群体遗传多样性越低[6]。本次研究测得9个黑山羊类群间核苷酸歧义度在0.287%~3.175%间变化,与李祥龙[7]和苗永旺[8]等对我国地方山羊遗传多样性及遗传背景研究相似。验证了David[9]、Gordon[10]研究结果。而总核苷酸歧义度为3.5%,表明群体内遗传变异大于群体间遗传变异,说明黑山羊群体遗传变异主要存在于种群内部;乐至黑山羊群体内遗传多样性指数较其他8个黑山羊群体大,说明乐至黑山羊群体内遗传变异程度较其他群体高,遗传纯度相对较低;米易黑山羊遗传多样性指数最小,说明在其自选过程中遗传变异程度较低,种质均匀性较好,遗传纯度较高,此结论与陈明华等[11]研究结果相一致,也与“大多数异交动植物遗传变异主要存在于群体内部,而自交生物遗传变异则主要存在于群体间”定律[12]相符合。也同金鑫燕等从地理范围、mtDNA和微卫星DNA等多个层次上对黑山羊群体遗传多样性研究结果基本一致[13-16]。也许是由于不同物种核内基因组遗传分化有不同机制或者是受不同因素影响所致。本研究认为,黑山羊由于分布地区自然环境和社会生态环境条件相似或不同,在自然选择与人工选择作用下,黑山羊群体遗传结构已经发生一些变异,部分群体形成比较丰富的遗传多样性,而部分群体变异程度较小。这是山羊养殖持续发展的物质基础,也可为四川山羊品系划分提供理论依据。
根据9个黑山羊类群遗传距离结合分析系统发育树,LZ与JT遗传距离比LZ与JC遗传距离更近,这与四川农业大学养羊研究室对乐至黑山羊mtDNA分子遗传特性研究结果相同。同时与王杰、郭鹏燕等对四川9个黑山羊群体DNA指纹分析[17]和RAPD标记研究得出结论相符。而9个类群黑山羊明显划分为A、B、C支系。由此可见,黑山羊起源并非单源,几种不同野生黑山羊群体参与黑山羊驯化,由于mtDNA只揭示母系参与驯化,但并不能确定参与驯化母系个数,本试验支持山羊多元起源观点[18-19]。
本研究首次对攀枝花本地黑山羊mtDNA D-loop区序列进行克隆测序,并对其基因结构进行初步分析,填补本研究领域空白。本研究可为从分子水平上探究黑山羊线粒体基因组遗传特点、物种进化关系以及物种多样性分析等提供科学参考。
[1] Mirol P M,Peral G P,Vega-Pla J L,et al.Phylogenetic relationships of argentinean creole horses and other south american and spanish breeds inferred from mitochondrial DNA sequences[J].Animal Genetics,2002,33:356-363.
[2] Sambrook J,Fritsch E H,Maiatis T.分子生物学实验指南[M].2版.金东雁,黎孟枫,侯云德等,译.北京:科学出版社,1998.
[3] 唐懿挺.西藏牦牛mtDNAcytb基因遗传多样性及ob基因原核表达[D].成都:西南民族大学,2011.
[4] Gojobori T,Ishii K,Nei M.Estimation of average number of nucle⁃otide substitutions when the rate of substitution varies with nucle⁃otide[J].Molecular Evolution,1982,18(6):414-422.
[5] 杨路存.几种鼢鼠mtDNA D-loop区序列多态性与系统发育关系研究[D].兰州:甘肃农业大学,2007.
[6] Zullo S,Sieu L C,Slightom J L,et a1.Mitochondrial D-loop se⁃quences are integrated in the rat nuclear genome[J].Journal of Molecular Biology,1991,221(4):1223-1235.
[7] 李祥龙.我国主要地方山羊品种遗传多样性及其起源分化研究[D].雅安:四川农业大学,1997.
[8] 苗永旺,叶绍辉,汪霞等.云南保种乳用圭山羊遗传多样性蛋白电泳研究[J].云南农业大学学报,1997,3(12):199-205.
[9] David E,MacHugh,Daniel G.Livestock genetic origins;Goats buck the trend[J].Proceedings of the National Academy of Scienc⁃es,2001,98:82-84.
[10] Gordon L,Ludovic G,Laurent E,et a1.Multiple maternal origins and weak phylogeographic structure in domestic goats[J].Proceed⁃ings of the National Academy of Sciences,2001,98:5927-5932.
[11] 王杰,陈明华,华太才让等.四川9个黑山羊品种(群体)微卫星DNA多态性研究[J].畜牧兽医学报,2006,37(11):1124-1129.
[12] 盛志廉,陈瑶生.数量遗传学[M].北京:科学出版社,1999.
[13] Gordon L,Ludovic G,Laurent E,et al.Multiple maternal origins and weak phylo geographic structure in domestic goats[J].PNAS,2001,98:5927-5932.
[14] 金鑫燕.10个山羊品种(群体)mtDNA D-loop序列多态性与成都麻羊MSTN基因克隆测序研究[D].成都:西南民族大学,2003.
[15] 马正花.四川8个山羊品种(群体)AFLP遗传多样性研究[D].成都:西南民族大学,2007.
[16] S.Sultana,Mannen H,S.Tsuji.Mitochondrial DNA diversity of Pakistani goats[J].Animal Genetics,2003,34:417-421.
[17] 王杰,郭鹏燕,王永,等.四川9个地方黑山羊品种(群体)DNA指纹分析[J].四川畜牧兽医,2005,32(10):27-28.
[18] Joshi M B,Rout P K,Mandal A K,et a1.Phylogeography and ori⁃gin of Indian domestic goats[J].Molecular Biology and Evolution,2004,21:454-462.
[19] Vila C,Seddon J,Ellegren H.Genes of domestic mammals aug⁃mented by baekerossing with wild ancestors[J].Trends in Genetics,2005(21):214-218.
