贵州省越冬区斑头雁种群动态与生境选择1)
2013-08-08周立晨罗祖奎牛俊英王天厚
周立晨 罗祖奎 牛俊英 王天厚
(华东师范大学,上海,200062) (凯里学院) (华东师范大学)
斑头雁(Anser indicus(Latham))是亚洲特有种,估计全球约7万只,越冬地主要分布在中国中部、西藏南部,印度,缅甸等区域,其中,2000只在云贵高原越冬[1]。斑头雁的血红蛋白具备强的氧亲和力结构,秋季迁徙时能够飞越氧气稀薄的喜马拉雅山脉顶峰,甚至能短时间的暴露在10 700 m的高空[2],其特殊的血红蛋白结构使之成为高海拔地区生存和繁衍的脊椎动物之一[3-4]。斑头雁在一些越冬区数量很不稳定[4-6],掌握该物种在越冬区的种群动态并分析种群数量变化的原因对该物种的保护有重要的意义。在2005年5月以前,对斑头雁的研究主要涉及到种群特征[7-8]、繁殖生态学[8-13]、血红蛋白晶体结构[2,14-17]、越冬行为[5]、食性[6]等多个方面。2005年5月中国青海湖发生了严重的野生水鸟感染高致病性禽流感事件之后,有关斑头雁的文献多与禽流感有关[1,18-31],仅有少量的文献涉及其生境选择[32]。
云贵高原是斑头雁的越冬地之一,贵州草海自然保护区及毗邻的杨湾桥水库(后文简称研究区域)是中国西南地区水禽的重要越冬区和中转站,斑头雁在研究区域的越冬时间一般在11月上旬—3月下旬。研究区域斑头雁数量波动显著[6],普遍认为中转站和越冬区生境丧失是斑头雁面临的最大威胁因素之一[4],对斑头雁生境选择的研究具有重要的科学价值[28]。多篇文献曾描述过斑头雁在越冬区均有明显的生境选择性[1,5-7],但由于研究的侧重点不同,很少文献量化斑头雁与生境之间的关系。而且,在云贵高原对于鸟类取食农作物没有实行生态补偿[33-34]或者补偿中断[5],人鸟矛盾没有协调好是研究区域偷猎现象屡禁不止的原因之一。因此,弄清越冬区斑头雁的种群动态、生境偏爱和食性选择,理论上有助于斑头雁越冬生态的研究,实践上有助于加强斑头雁及其生境的保护管理。本研究目的有3点:①比较研究区域斑头雁数量在不同生境中的差异性,找出斑头雁偏爱的生境;②探究蔬菜地中斑头雁数量与植被盖度的线性关系,为制定生态补偿措施提供依据;③探讨自然环境中影响斑头雁数量和分布的关键因子,为该物种的保护提供理论依据。
1 研究区概况
研究区域位于贵州省毕节地区威宁县城西南侧,属亚热带半湿润季风气候,海拔为2 170~2 386 m。年降水量为950.9 mm,冬、春季降水总量不足全年的12%。最高平均温度在7月为17.7℃,最低平均温度在1月为1.9℃,年平均温度10.6℃。研究区域包括草海国家级自然保护区(26°47'32″~26°52'52″N,104°10'16″~104°20'40″E)和杨湾桥水库(26°48'30″~26°52'30″N,104°07'30″~104°10'20″E)2个高原淡水湖泊,是中国西南地区水禽的重要越冬区和中转站(图1)。其中,草海自然保护区是云贵高原最大的天然淡水湖泊,保护区总面积为96 km2,冬季水面积 25 km2,冬季水深为 1.35 ~ 2.80 m,整个冬季水位呈下降趋势,草海保护区以保护黑颈鹤(Grus nigricollis)为代表的珍稀鸟类和高寒湿地生态系统为主。杨湾桥水库是1958年修建的人工蓄水湖,冬季水面积4.85 km2,平均水深超过7 m。

图1 研究区域地理位置
2 研究方法
2.1 鸟类调查
2010和2011年冬季(12月—翌年2月)以及2011年春季(3月)对研究区域的斑头雁进行野外调查。将生境分为农田、蔬菜地、草甸、水域和树林5种类型。除了水域以外,其他生境呈小斑块交错分布的景观格局,根据生境分布状况,所有的样线设置为长400 m、宽100 m,所有样线之间间隔超过100 m。选择晴天用双筒望远镜(型号:BD42 Series Kowa 8×),以1 km·h-1的速度记录单侧100 m内的斑头雁数量。累计样线数209条,其中冬季173条、春季36条。样线随机选择并且单个季节内所有样线的调查不重复。
2.2 生境调查
记录每条样线内斑头雁数量的同时记录生境特征,研究区域生境类型及其特征的描述见表1。为探究蔬菜地中斑头雁数量与植被盖度的关系,用测距仪(型号:Bushnell Pro 500)量取每条样线内萝卜菜、白菜、紫花苜蓿以及其他植被的占地面积。
2.3 数据处理
比较同一生境中斑头雁冬、春季之间的数量差异以及比较同一种蔬菜冬、春季之间的盖度差异时,数据不满足正态分布的要求,选用Mann-Whitney U Test检验。比较同一季节5种生境之间斑头雁数量差异时,数据不满足正态分布的要求,选用Kruskal-Wallis Test检验。用多元回归(Stepwise法)分析蔬菜地中斑头雁数量与植被的线性关系时,以斑头雁数量为因变量,以萝卜菜盖度、白菜盖度、紫花苜蓿盖度、其他植被盖度4组变量为自变量,4组自变量均满足正态分布的要求(P=0.062~0.326,One-Sample Kolmogorov-Smirnov test);4组自变量在冬季(r=-0.210 ~0.099,P=0.283 ~0.975)和春季(r=-0.183 ~ 0.168,P=0.532 ~ 0.933)两两之间的Pearson相关性均较弱,表明多元回归分析时数据之间共线性较弱。所有统计分析均在 SPSS 16.0(SPSS Inc.)上运行。
3 结果与分析
3.1 斑头雁季节动态
斑头雁数量在草甸冬季显著高于春季(U=38.500,P=0.019,Mann-Whitney U Test),在树林春季极显著高于冬季(U=98.000,P=0.002),但在农田(U=118.500,P=0.140)、蔬菜地(U=165.000,P=0.343)和水域(U=356.000,P=0.834)斑头雁数量冬、春季之间无显著差异(表2)。
3.2 斑头雁生境选择
冬季不同生境之间斑头雁数量差异极显著(χ24=20.222,P<0.001,Kruskal-wallis Test),平均值从高到低依次为:水域、草甸、蔬菜地、农田、树林。春季不同生境之间斑头雁数量差异显著(χ24=9.994,P=0.041),平均值从高到低依次为:草甸、农田、蔬菜地、水域、树林(表2)。不同生境中斑头雁数量变异系数由小到大依次为:农田、水域、蔬菜地、草甸、树林(表3)。

表1 研究区域斑头雁生境类型及其特征
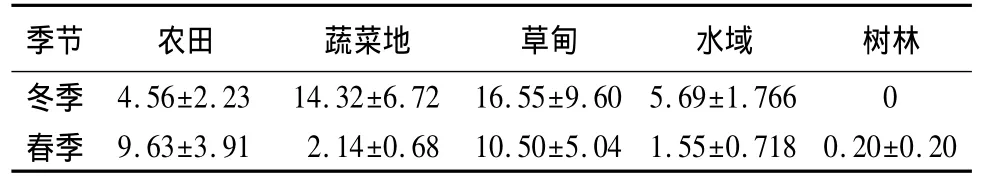
表2 斑头雁数量在不同季节、不同生境中的差异性只·样线-1
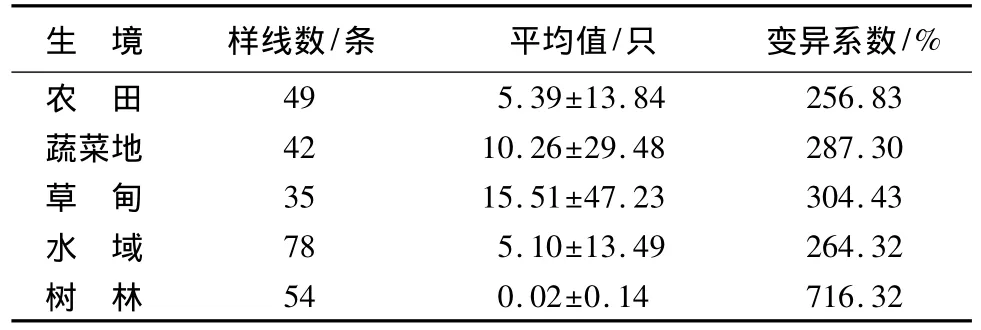
表3 不同生境中斑头雁数量变异系数
3.3 蔬菜地中斑头雁数量与植被盖度的关系
3.3.1 蔬菜地中植被季节变化
萝卜菜盖度冬季显著高于春季(U=113.500,P=0.023,Mann-Whitney U Test),白菜盖度(U=172.000,P=0.498)、紫花苜蓿盖度(U=172.500,P=0.503)、其它植被盖度(U=165.000,P=0.296)冬、春季之间无显著差异(表4)。
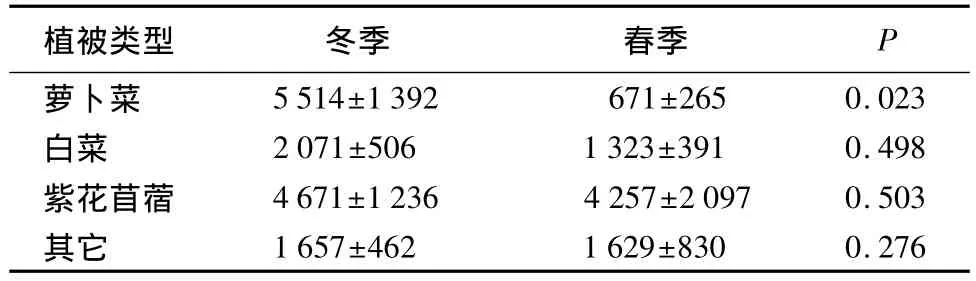
表4 蔬菜地不同植被冬、春季盖度 m2·样线-1
3.3.2 斑头雁数量与蔬菜地植被盖度的关系
多元回归(Stepwise法)分析显示(表5):在冬季,斑头雁的数量与萝卜菜和紫花苜蓿的盖度呈极显著正相关性(R2=0.400,P=0.002),但白菜和其他植被盖度为剔除变量(P>0.05);在春季,斑头雁的数量与紫花苜蓿的盖度呈极显著正相关性(R2=0.630,P=0.001),但萝卜、白菜和其他植被盖度为剔除变量。
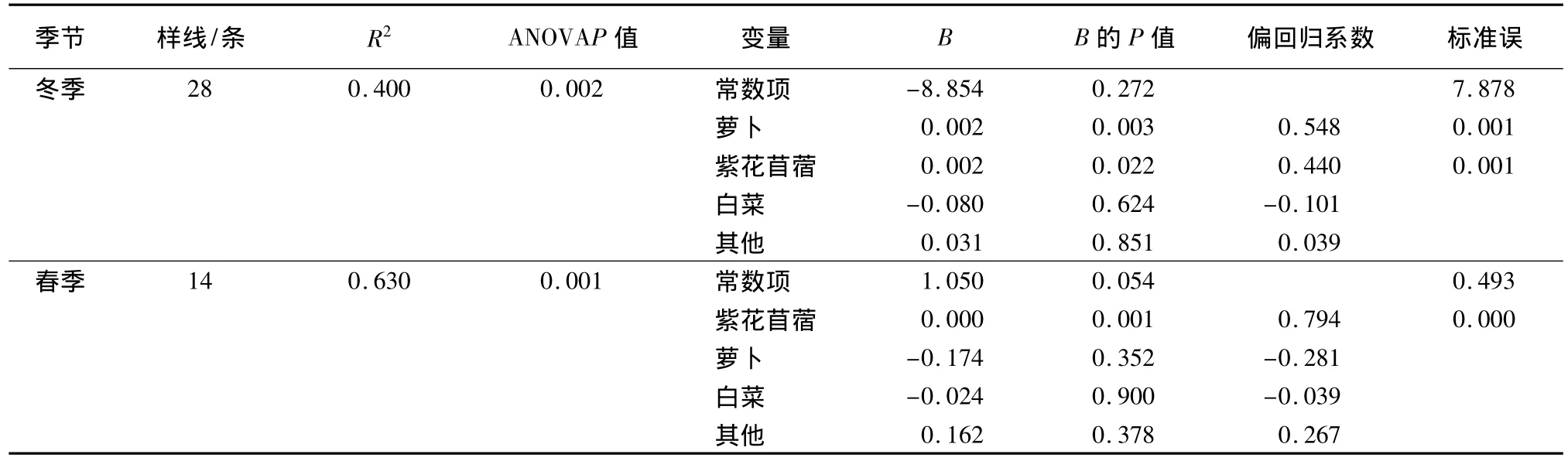
表5 斑头雁数量与蔬菜地中不同种类的植被盖度的回归关系(Stepwise法)
4 结论与讨论
4.1 斑头雁集群行为
对云南拉市海越冬期斑头雁行为观察发现,斑头雁无论取食还是歇息,都喜集群,特别是在麦地活动时喜集大群[5]。本研究区域生境异质性高,斑头雁常在草甸和萝卜菜地生境中集大群,斑头雁的集群使其种群数量在每个样方中差异很大,导致各生境中斑头雁数量的变异系数均较大(表3),同时,也导致数据分析时数据分布方差不齐和不符合正态分布。斑头雁大量的个体聚集增加了病原体感染的风险,形成一种生存压力[22]。有研究表明,斑头雁无论是白天活动[1,5]还是夜栖[5]常与赤麻鸭(Tadorna ferruginea)混群。笔者观察发现,在草甸和浅水区斑头雁、赤麻鸭和黑颈鹤(Grus nigricollis)经常混群活动,而且这3种鸟类在草海的阳关山和胡叶林区域也掺杂夜栖。
4.2 斑头雁季节数量变化
草海自然保护区[6]及其邻近拉市海[5]斑头雁在3月上旬开始迁离,但一直持续到3月下旬才全部迁完,迁徙持续时间近一个月。斑头雁的迁离使其春季数量比冬季对应生境中的数量要少,虽然在树林中春季的数量比冬季的高,但是,整个调查中树林样线中总共仅记录到2只斑头雁,所以,在树林中斑头雁春季的数量比冬季的高只能归为一种偶然因素(表2)。
4.3 斑头雁越冬生境偏爱与食物选择
虽然越冬期间的斑头雁能在浅水、农田、岸边泥滩地等多种生境中觅食[5],但由于斑头雁是食草为主的水禽[9-10],对食物有选择性[6],因此,表现出对生境有明显的选择性[1],越冬期间高寒生态系统中的斑头雁喜好湖泊河流周边的湿地沼泽类生境[7]。研究表明,冬季和春季不同生境之间斑头雁数量均存在显著的差异(表2)。斑头雁对生境利用的差异性不仅表现在对不同生境的选择上[6],还表现在对同类生境不同时间段的选择上[5],比如:草甸中斑头雁的数量冬季显著高于春季(表2)。禾本科植物是斑头雁主要食物,草海越冬的斑头雁粪便中禾本科植物的检出率为65.0%[6]。对拉市海越冬的斑头雁观察发现,冬季斑头雁不选择的河滩泥地,到了冬末春初(3月上旬)由于河滩泥地长出禾本科植物的草芽会使其成为斑头雁重新选择的生境[5]。1998年1月对草海自然保护区的斑头雁进行统计,记录到75%的个体出现在草甸生境[6]。研究显示,草甸生境中斑头雁的平均数量冬、春季在5类生境中均较高(表2),这可能与禾本科植物主要分布在草甸生境有关。但是,观察发现,斑头雁在农田和蔬菜地中多数时间在觅食,而在草甸和水域生境多为休息,比如:在草甸生境中斑头雁在16:30—19:00高频率取食,而在其他时间多在休息,斑头雁选择水域和草甸生境休息可能与人为活动少有关。虽然斑头雁是游禽,但越冬期间其食物主要分布在陆地而不是水体中[5],特别是斑头雁很少到深水区活动[5-6],笔者的观察结果与此一致(表2)。
紫花苜蓿在斑头雁粪便中检出的频率不超过6%,但占粪便中农作物总检出率的90%以上[6]。研究表明,冬季蔬菜地中斑头雁的数量与萝卜和紫花苜蓿的盖度呈强的正相关,春季蔬菜地中斑头雁的数量仅与紫花苜蓿盖度呈强的正相关(表5),野外观察发现,冬季萝卜菜和紫花苜蓿的叶都是斑头雁主要取食对象,而较少取食白菜、大蒜等其他植物。在研究区域,萝卜菜的收割主要在1月下旬,导致冬、春季蔬菜地萝卜菜的盖度差异极显著(表4)。春季即使有少量的萝卜没有被收获,萝卜的叶也已经被赤麻鸭和斑头雁等鸟类取食殆尽,甚至露出地表的部分萝卜也被取食过,也就是说春季的萝卜菜已经没有斑头雁可以食用的部分,这可能是蔬菜地中斑头雁与萝卜菜盖度只在冬季呈显著正相关性,而春季相关性不显著的原因。在整个越冬期间斑头雁均取食紫花苜蓿[6],这种植物是当地农民为改良土壤结构而种植的一种绿肥,牛羊过食紫花苜蓿容易引发瘤胃胀气而死亡,但斑头雁可以连续的取食而无生命危险值得研究。
4.4 物种保护与生态补偿
在拉市海用稻草人等方式驱赶小麦地中的斑头雁,但并不能阻止斑头雁到麦地取食麦苗,而鸟类损害庄稼多年未得到政策性的生态补偿,人鸟矛盾突出,驱赶鸟的事件经常发生[5]。对草海斑头雁取食农作物的解决方法,有生态学者建议购买靠近沼泽地的部分农地种植斑头雁喜欢的植物,或在寒冷的月份购买部分农地上的农作物[6]。笔者发现,不只是斑头雁取食农作物,赤麻鸭、黑颈鹤、灰鹤等均为常见的取食农作物的鸟类,前期的生态学者提出实施生态补偿措施有利于草海自然保护区的人鸟和谐[33-34]。
[1] 刘冬平,张国钢,钱法文,等.西藏雅鲁藏布江中游斑头雁的越冬种群数量、分布和活动区[J].生态学报,2010,30(15):4173-4179.
[2] 梁宇和,查正,华子千,等.空间和地面条件下斑头雁血红蛋白晶体结构的比较研究[J].北京大学学报:自然科学版,2001,37(4):483-489.
[3] 张耀南,郝美玉,雷富民,等.环青海湖斑头雁种群数量动态模拟及趋势分析[J].动物学研究,2009,30(5):578-584.
[4] Köppen U,Yakovlev A P,Barth R,et al.Seasonal migrations of four individual bar-headed geese Anser indicus from Kyrgyzstan followed by satellite telemetry[J].Journal Fur Ornithologie,2010,151:703-712.
[5] 刘宁,李德品.拉市海越冬末期斑头雁行为的初步研究[J].西部林业科学,2004,33(4):69-74,89.
[6] 李凤山,聂卉,叶长虹.贵州草海斑头雁的冬季食性分析[J].动物学杂志,1998,33(4):29-33.
[7] 宋延龄,Bishop M A,仓曲卓玛.西藏雅鲁藏布江中游地区斑头雁越冬种群数量和分布[J].动物学杂志,1994,29(2):27-30.
[8] Prins H H T,van Wieren S E.Number,population structure and habitat use of bar-headed geese Anser indicus in Ladakh(India)during the brood-rearing period[J].Acta Zoologica Sinica,2004,50(5):738-744.
[9] 冼耀华.青海湖地区斑头雁繁殖习性的初步观察[J].动物学杂志,1964,6(1):12-14.
[10] 王侠.斑头雁繁殖习性的观察[J].野生动物,1981(3):29-31.
[11] Lamprecht J.Female reproductive strategies in bar-headed geese(Anser indicus):Why are geese monogamous?[J].Behavioral Ecology and Sociobiology,1987,21:297-305.
[12] Schneider J,Lamprecht J.The importance of biparental care in a precocial,monogamous bird,the bar-headed goose(Anser indicus)[J].Behavioral Ecology and Sociobiology,1990,27:415-419.
[13] 马鸣,才代.天山巴音布鲁克斑头雁巢的聚集分布及其繁殖生态[J].应用生态学报,1997,8(3):287-290.
[14] 卢光莹,毛春虹,卫新成,等.斑头雁(Anser indicus)高铁血红蛋白的晶体生长和初步的X射线衍射研究[J].生物物理学报,1989,5(3):259-262.
[15] 华子千,朱红,卫新成,等.斑头雁(Anser indicus)氧合血红蛋白的结晶和X射线的初步分析[J].生物物理学报,1990,6(2):221-223.
[16] Wang Huanchen,Liang Yuhe,Zhu Jiapeng,et al.Crystallization and preliminary crystallographic studies of bar-headed goose fluoromethaemoglobin with inositol hexaphosphate[J].Acta Crystallographica Section D-Biological Crystallography,2000,56(9):1183-1184.
[17] Liu Xiaozhou,Li Songlin,Jing Hua,et al.Avian haemoglobins and structural basis of high affinity for oxygen:structure of barheaded goose aquomet haemoglobin[J].Acta Crystallographica Section D-Biological Crystallography,2001,57:775-783.
[18] 白杉.候鸟导致全球禽流感危机[J].山西农业,2006(1):27-29.
[19] 郑杰,何玉邦.对青海湖区野生鸟类禽流感疫情防控的几点思考[J].野生动物,2006,27(5):19-21.
[20] Chen H X,Shen H G,Li X L,et al.Seroprevalance and identification of influenza a virus infection from migratory wild waterfowl in China(2004-2005)[J].Journal of Veterinary Medicine Series B,2006,53(4):166-170.
[21] Spackman E,Swayne D E,Gilbert M,et al.Characterization of low pathogenicity avian influenza viruses isolated from wild birds in Mongolia 2005 through 2007[J].Virology Journal,2009,6:190-198.
[22] 朱筱佳,李来兴,杨乐,等.青海湖鸟岛斑头雁种群对H5N1亚型禽流感病毒的免疫状况[J].动物学研究,2009,30(4):406-410.
[23] Sivay M V,Silko N Y,Sharshov K A,et al.The role of wild goose(Anser)populations of Russia and the Tibet Plateau in the spread of the avian influenza virus[J].Chinese Birds,2011,2(3):140-146.
[24] 楚国忠,侯韵秋,张国钢,等.卫星跟踪青海湖繁殖水鸟的迁徙[J].自然杂志,2008,30(2):84-89.
[25] 刘冬平,张国钢,江红星,等.青海湖斑头雁(Anser indicus)繁殖期的活动性、栖息地利用及其与人的关系[J].生态学报,2008,28(11):5201-5208.
[26] Prosser D J,Takekawa J Y,Newman S H,et al.Satellitemarked waterfowl reveal migratory connection between H5N1 outbreak areas in China and Mongolia[J].Ibis,2009,151:568-576.
[27] Gilbert M,Newman S H,Takekawa J Y,et al.Flying over an infected landscape:distribution of highly pathogenic avian influenza H5N1 risk in South Asia and satellite tracking of wild waterfowl[J].EcoHealth,2010,7:448-458.
[28] 郝美玉,张耀南,雷富民,等.基于数字地形模型的斑头雁潜在生境分析[J].动物学杂志,2010,45(1):35-42.
[29] 张国钢,刘冬平,钱法文,等.西藏夯错水鸟多样性及斑头雁繁殖活动区的变化[J].生态学报,2010,30(2):395-400.
[30] Iverson S A,Gavrilov A,Katzner T E,et al.Migratory movements of waterfowl in Central Asia and avian influenza emergence:sporadic transmission of H5N1 from east to west[J].I-bis,2011,153:279-292.
[31] Cui Peng,Hou Yuansheng,Tang Mingjie,et al.Movement patterns of bar-headed geese Anser indicus during breeding and postbreeding periods at Qinghai Lake,China[J].Journal Fur Ornithologie,2011,152:83-92.
[32] 侯元生,崔鹏,星智,等.青海湖主要集群繁殖鸟类巢区分布及其生境特征[J].野生动物,2010,31(3):131-134.
[33] 罗祖奎,李性苑,张文华,等.草海自然保护区春季鸟类群落结构及现存生物量[J].湖北大学学报:自然科学版,2011,33(4):408-412,417.
[34] 罗祖奎,刘文,李振吉,等.贵州草海冬季鸟类群落特征[J].华东师范大学学报:自然科学版,2012(4):102-111.
