浙江三门湾沉积记录:U k37表层海水温度和浮游植物群落 变化与El Niño/La Niña的关联
2013-07-13赵庆英陈荣华卢冰叶新荣胡锡钢
赵庆英,陈荣华,卢冰,叶新荣,胡锡钢
1. 国家海洋局第二海洋研究所,国家海洋局第二海洋研究所工程海洋学重点实验室,浙江 杭州 310012;
2. 国家海洋局第二海洋研究所,国家海洋局海洋生态系统和生物地球化学重点实验室,浙江 杭州 310012
厄尔尼诺(El Niño)和拉尼娜(La Niña)是2种发生在海洋中的自然灾害现象,其显著特征是赤道中东太平洋海域的海水出现异常增温(El Niño现象)或降温(La Niña现象),它们经常交替循环发生。El Niño / La Niña的生成、发展和消亡可导致世界沿海各地发生程度不一的自然灾害,也给全球海洋环境和生物群落带来显著影响[1-5]。El Niño/La Niña对我国气候也有显著的影响。国内外对El Niño/La Niña的研究已取得许多重要成果,然而同时,限于模式的不完善,迄今对El Niño变化机理与发展趋势的解译尚缺乏一致性,所使用的耦合模式在模拟El Niño变率变化方面的不确定性依然很大。为了应对新的挑战和提高对El Niño/La Niña 的预测能力,学术界不仅对海气耦合模型提出了更为苛刻的要求,并试图寻找新的研究途经和解决问题的钥匙。于是,利用不同历史时段El Niño/La Niña事件的自然记录来研究和揭示其现象特征与本质[6-9],进行“过去的El Niño/La Niña”关键变化过程的分析研究,将成为最直接有效和具有开拓性的工作。本研究可为浙江三门湾海域约180年来生物标志物重建浮游植物种群结构提供依据,并与现场调查资料相符。
本文研究选择浙江三门湾内近岸区域的沉积柱样。三门湾为半封闭海湾,沉积环境稳定,沉积速率高,又是季风作用的地区,适宜生物标志物的完整保存,并且有高分辨率的沉积地层记录。本文利用长链烯酮不饱和指数Uk37与温度的特性,直接辨识和建立近180年来El Niño/La Niña现象,捕捉分子地层学记录中可能的历史相似型的信号,并利用三门湾现场实测浮游植物数据来分析群落结构变化与El Niño的关联程度;同时选取浮游植物标志物对海洋初级生产力进行研究,利用菜子甾醇(硅藻Diatom)、甲藻甾醇(沟鞭藻Dinoflagellates)和C37直链烯酮(颗石藻Emiliania huxleyi)之和作为初级生产力的总量,浮游植物标志物的比值(菜子甾醇/甲藻甾醇)指示浮游植物种群结构的变化[10-12],将这些指标进行综合研究,试图给出新近频频发生的El Niño/La Niña事件对浙江省沿岸海域浮游植物群落的具体影响。
1 样品采集与分析方法
1.1 样品采集
H30柱状样为重力活塞样,于1995年4月采于三门湾猫头嘴附近(29°18′ 50E′′,121°64′ 00′′ N,长度约为250 cm),柱样分割间距一般为1.0 cm;26站表层沉积物样品2次采于1997年8月和1998年9月,其余沉积物样品采于2002年8月和2003年5月;S1~S18站的三门湾四季浮游动物调查采于2002年8月和11月,2003年2月和5月,站位见图1。同海域对比资料来自《三门湾生态环境调查综合报告》(2005年/2006年)4个航次的调查成果。

图1 三门湾样品采集站位图 Fig.1 Map of the in Sanmen Bay with the sampling site
1.2 分析方法
样品干燥与研磨后,加入二氯甲烷和甲醇混合溶剂(体积比3∶1),加入内标20 µL异30烷(r-C30)、24氘烷(n-C24D50)和19醇(C19H39OH)索氏提取48 h(抽提水浴温度控制在62 ℃左右、回流速度为4~5次/h),获得总的可提取有机质,加入活化铜片,放置过夜脱硫,提取液经旋转蒸发浓缩。再加入含60 g/L KOH的甲醇溶液,进行碱水解除去酸性组分;离心机分离,用正已烷提取4次,上清液经旋转蒸发浓缩,最后用氮气吹干。用正已烷转移经硅胶层析柱分离(硅胶经抽提后在180 ℃热活化12 h,待冷至室温时再加入其重量的3%的去离子水去活化,放置过夜使之达到平衡;氧化铝在250 ℃热活化12 h)。用正已烷淋洗收集烷烃组分;用二氯甲烷(加50 g/L甲醇)淋洗收集醇类、烯酮组分。将上述两部分组分用氮气吹干,分别转移至1 mL刻度管内,其中烷烃直接上机GC分析。醇类、烯酮组分加入二氯甲烷和双(三甲基硅烷基)三氟乙酰胺(BSTFA)衍生化,转变为三甲基硅醚衍生物(TMS—醚),以备进行GC分析。图2为沉积柱样中生物标志物典型气相色谱图。

图2 (a)柱样中生物标志物典型气相色谱图和(b)C37:2Me质谱图 Fig.2 (a)Gas chromatograms of phytoplankton biomarker and (b)mass spectrometry of C37:2Me
1.3 仪器条件
醇类、烯酮:气相色谱(Agilent 5890)分析,弹性石英毛细管柱(D8-5、30 m×0.25 mm)。升温程序:初始温度80 ℃,升温速度25 ℃/min升至200 ℃, 4 ℃/min升至250 ℃, 1.8 ℃/min升至300 ℃,5 ℃/min升至310 ℃并保持5 min。所用载气:氢气,流速1.2 mL/min。含量由其峰面积与内标面积对比计算得到。数据的相对偏差<5%,由于有些样品长链烯酮含量很低,没有做回收率。长链烯酮一般在m/z 57、67、71、82、96、278、486、528、530、542、556和558等附近有特征碎片峰。
1.4 浮游植物观察、鉴定
根据不同的样品选择用中性甲醛、戊二醛或碘液固定。样品经浓缩后用日本生产Olympus-VANOX-AHB: LB-2万用显微镜观察、鉴定和计数,所有操作均按《海洋调查规范》进行。初级生产力采取表层水样进行培养,水样加入培养瓶前,经280 µm孔宽的筛绢预过滤,以除去大多数浮游动物。初级生产力的粒度分级测定水样依次经孔宽20 µm筛绢、孔径2 µm的Nuclepor 滤膜和Whatman GF/F玻璃纤维滤膜过滤,分离小型、微型和微微型光合浮游生物。
2 结果与讨论
2.1 年代地层标定
地质时间是我们认识一切地质作用、过程和产物的重要参数。210Pb是天然放射性U-Th系的一个子体,具有22.3 a半衰期,是百年尺度测年的一个极好的核素,现被广泛应用于海洋沉积物的测年。取12个层段样品,采用210Pb方法进行测年;用S = λ1×Z/In ( Co/Cz ) 计算沉积速率。式中:S为沉积速率,单位:cm/a;λ1为210Pb衰变常数为0.03114/a;Z为深度,单位:cm;Co、Cz分别为表层与深度Z层。分析结果表明,H30柱样210Pb剩余比活度的时间跨度为1820—1995年(始于约175年前),大多层位沉积速率为1.0 cm/a(图3和表1);该测年为高分辨率的气候研究提供保证。2个表层样(26站和05站)依据当年采集样品时间定年为2005年和1997年。
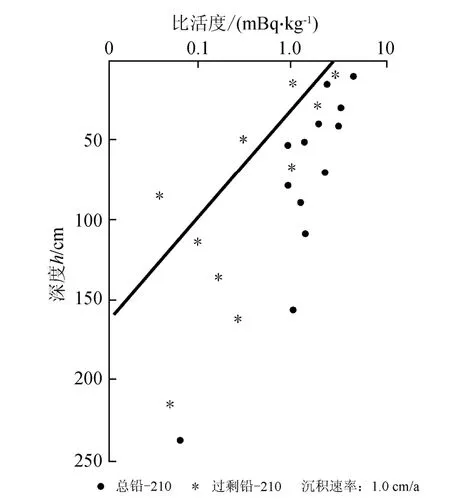
图3 三门湾H30柱样210Pb比活度垂向分布 Fig.3 Downcore 210Pb specific activities in the core H30 from Sanmen Bay
2.2 长链烯酮不饱和比值Uk37与表层海水温度
H30柱样中长链烯酮化合物分别为甲基酮化合物(C37:3Me、C37:2Me、C38:3Me、C38:2Me)和乙基酮化合物(C38:3Et、C38:2Et),其中以C37:2Me含量最高,其次是C38:2Et,主要来源于定鞭藻门颗石藻纲钙质鞭毛藻科(一种个体微小的单细胞真核藻,其特殊之处在于藻体外表附着有结构精巧的球石粒构成的球石层,主要成分是CaCO3,主要生长在海水上部透光层,对光线强度和营养物质丰富程度反应极为灵敏)。在三门湾H30柱样中的长链烯酮含量很低,可能与颗石藻是海洋生物的第一食物链并在沉积过程中被溶解有关。
异常的海洋与气候所造成的海水增温或降温,必然会影响海洋藻类(颗石藻)合成长链烯酮化合物,长链烯酮不饱和指数Uk37这种信息会在沉积速率高、扰动小的深海得以完整地保存,Uk37与表层海水温度有很好的线性关系[13-15]。本次研究应用Pelejero等(1997)南海Uk37校正公式Uk37=0.031SST+0.092[16],Uk37波动范围在0.555~0.642,H30柱样中长链烯酮不饱和指数波动范围在15.19~17.61 ℃(见表1)。研究结果表明,三门湾近180年以来表层海水温度呈现上升趋势,与全球气温变暖的趋势一致。该海湾在2002年实测年平均水温为20.55 ℃(春季19.21~23.57 ℃;夏季27.21~29.74 ℃;秋季17.21~18.14 ℃;冬季9.47~10.75 ℃)[17],因此Uk37估算温度要低于实际年均SST,而与秋季表层海水温度接近,可能秋季是颗石藻群落旺发季节。
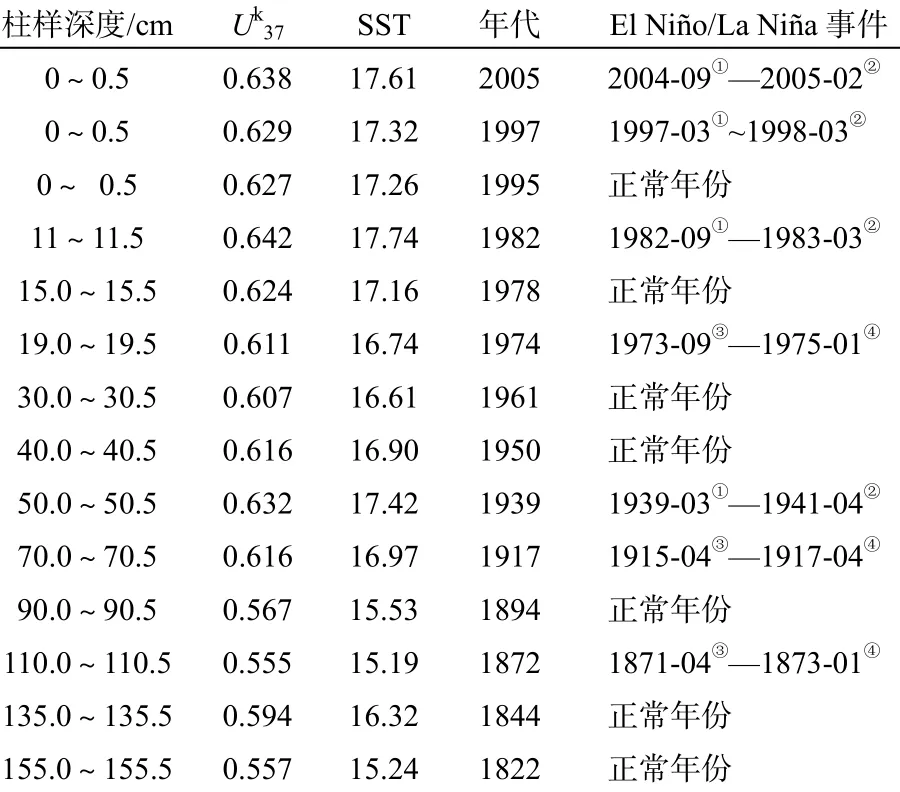
表1 三门湾H30柱样210Pb地质年龄及表层样年代 Table 1 210Pb dating for the sediment core H30 and surface sediment samples from Sanmen Bay
2.3 估算的表层海水温度与El Niño/La Niña的遥相关
本文将El Niño年、La Niña年、正常年份划分为3个等级来研究,El Niño比La Niña的生命周期短,但强度大,盛期明显;而拉尼娜持续时间较长。表1可见,从1822年到2005年,三门湾H30柱样沉积中记录的厄尔尼诺有4次,拉尼娜3次,正常年份7个。Uk37估算温度清晰捕捉了该海域的温度变化、并在El Niño/La Niña年份中表现出明显的增/降幅度。
2004年9月El Niño在太平洋中部赤道水域形成,但比较温和,这一过程持续到2005年2月结束,2005年的Uk37估算温度为17.61 ℃;1997年3月至1998年6月是100年来最强烈的一次El Niño事件,1997年夏季,正距平已经移到了中东太平洋,海表温度剧烈上升,形成一次异常猛烈的气候事件(1997年的Uk37估算温度为17.32 ℃);1982—1983年也是一次较强的El Niño年事件(1982年的Uk37估算温度为17.74 ℃);同样,在El Niño状态下的1939年,Uk37估算温度为17.42 ℃。作为对比,三门湾H30柱样记录中7个正常年份(1995年、1978年、1961年、1950年、1894年、1844年和1822年),Uk37估算温度分别为17.26、17.16、16.61、16.90、15.53、16.32、15.24 ℃。由此可见,以厄尔尼诺年的Uk37估算温度总体高于正常年份。分析La Niña事件时,我们发现沉积地层中Uk37估算的温度记录相对于La Niña年不太明显,只有少数拉尼娜了事件留下痕迹:1973—1975年间发生2次强La Niña事件,1974年地层记录的Uk37估算温度为16.74 ℃;1915—1917年为最强冷事件,1917年地层记录的Uk37估算温度为16.97 ℃;1872年地层记录的Uk37估算温度为15.19 ℃。这表明三门湾SST对于La Niña事件的响应可能并不明显。
2.4 地层记录中浮游植物群落变化与El Niño/La Niña的关联
浮游植物驱动了整个海洋生态群落以及其中的生命,浮游植物对环境变化敏感,通过对沉积物中浮游植物的生产量和群落组成变化研究,将有利于我们了解浮游植物与气候变化的对应关系。本文选取浮游植物标志物对海洋初级生产力群落进行研究。浮游植物标志物含量不但可以作为单种浮游植物含量的参数,浮游植物标志物的比值(菜子甾醇/甲藻甾醇)也可以指示浮游植物种群结构的变化,利用菜子甾醇(硅藻)、甲藻甾醇(沟鞭藻)和C37直链烯酮(颗石藻)组成初级生产力群落,将它们之和作为初级生产力的总量,这些指标结合起来研究,在一定程度上可以反映浮游植物群落结构的变化,及对El Niño /La Niña变化作出的响应。
从表2可看出,三门湾H30柱样中的3种生物标志物总含量(菜子甾醇+甲藻甾醇+长链烯酮)的变化范围为710.7~6998.5 ng·g-1。生物标志物估算的总初级生产力在近180年的时间里,总体呈上升趋势,单一浮游植物标志物菜子甾醇(硅藻)、甲藻甾醇(甲藻)和C37直链烯酮(颗石藻)的含量变化各在523.1~5955.7、180.5~1132.9、7.1~44.1 ng·g-1,显示出沉积物中硅藻相对含量较高,甲藻次之,颗石藻所占比例很低,但是有离岸越远比例升高的趋势。从地质历史的角度来看,浮游植物群落的物种组成、丰度有很大的变化,但一直以硅藻占优势,所占比例呈上升趋势。单一浮游植物变化曲线表明了种群结构具有明显变化,主要表现在硅藻和甲藻相对含量变化上,估算的硅藻在67.65%~87.3%、甲藻在12.2%~31.7%、颗石藻在0.5%~1.3%;硅藻/甲藻比值在2.1~7.2。

表2 三门湾柱样沉积物中生物标志物含量变化和估算的单种藻类相对含量 Table 2 Biomarker content changes and relative content of single species of algae in the sediment core from Sanmen Bay
三门湾H30柱样沉积地层记录的El Niño事件,2005年(2004-09—2005-02)、1997年(1997-03— 1998-03)、1982年(1982-09—1983-03)、1939年(1939-03—1941-04),利用浮游植物标志物估算的初级生产力含量相对较高为4998.5~5406.0 ng·g-1,菜子甾醇估算的硅藻在82.0%~87.3%,甲藻甾醇估算的甲藻为12.2%~17.3%,硅藻/甲藻比值为4.7~7.2,对以上浮游植物群落结构进行比较,三门湾海洋生态对海洋环境变化的敏感性,浮游植物的生产力在El Niño期间有所提高;沉积地层记录的La Niña事件,1974年(1973-09—1975-01)、1917年(1915-04—1917-04)、1872年(1871-04— 1873-01),利用浮游植物标志物估算的初级生产力含量为(710.7~1482.9 ng/g-1),菜子甾醇估算的硅藻在67.6%~73.6%,甲藻甾醇估算的甲藻为25.4%~31.7%,硅藻/甲藻比值为2.1~2.9。对以上El Niño/La Niña期间的浮游植物群落结构进行比较,发现El Niño/La Niña事件影响到不同浮游植物单体增减,特别是硅藻与甲藻的关系与El Niño/La Niña事件有密切关联。在这里需要提出的是,在沉积地层记录中Uk37估算温度在La Niña冷事件中,只有少数地层留下了记录;而浮游植物标志物估算的藻类在La Niña期间得到了很好的反映,这表明三门湾海域浮游植物群落盛衰不仅仅受控于温度。
2.5 (实测)浮游植物群落变化对El Niño/La Niña的响应
2002/2003年El Niño事件(2002-04— 2003-03),在4月份赤道东太平洋Niño3区(美国国家气候分析中心CAC所用的指标区[28])的SST距平(SSTA)为0.16 ℃,在5月份爆发(为0.19 ℃),到6月升为0.68 ℃;El Niño在11月达到峰值,2002年12月为1.41 ℃,热带大气、海洋状况仍为El Niño盛期,2003年1月SST距平迅速减弱,这次El Niño在2003年3月结束,现场生物实测时间段在(2002-08、2002-11、2003-02和2003-05) 正好处于El Niño过程中,共鉴定出浮游植物5门133种,硅藻和甲藻是调查区的主要浮游植物类群,浮游植物生态类型多为温带近岸种,少数为暖水种或大洋种。
随着当年气温升高、浮游植物生长率明显提高,浮游植物的生长率与其现存量(叶绿素a)表现出一定的相关性,El Niño年提高了浮游植物生产率,可能是由于增加的降雨以及径流带来了比以往更多的养分,使得浮游植物蓬发、细胞丰度达到1 516.61(×103·dm-3),但浮游植物种类减少到85种,为周年最低,同时高温高盐暖水性浮游植物大量出现,浮游植物与营养盐状况也有很好的一致性[17];秋季剧减到9.11(×103·dm-3),当浮游植物遇到各种各样的环境胁迫时,如在水华发生过程中浮游植物的生长逐渐受到限制,最后导致浮游植物自动死亡。
2006年是一次弱El Niño现象,当年4月在赤道中东太平洋初步形成,8月赤道中东太平洋NINO综合区海温指数为0.6,表明热带海洋大气进入El Niño状态,本文讨论的春季生物现场调查在4月初,通过大气遥相关来传播其影响[18],东亚大气环流对El Niño事件大气响应具有滞后效应。现场生物实测时间段在2005年(8月、11月、2月和5月)和2006年(7月、10月、1月和4月)之间,故而2005—2006年可作为正常年份来讨论。当年三门湾共鉴定出浮游植物6门56属146种,其中硅藻38属115种(占78.8%),甲藻12属25种(占17.1%),其他蓝藻、金藻和绿藻等占4.1%。
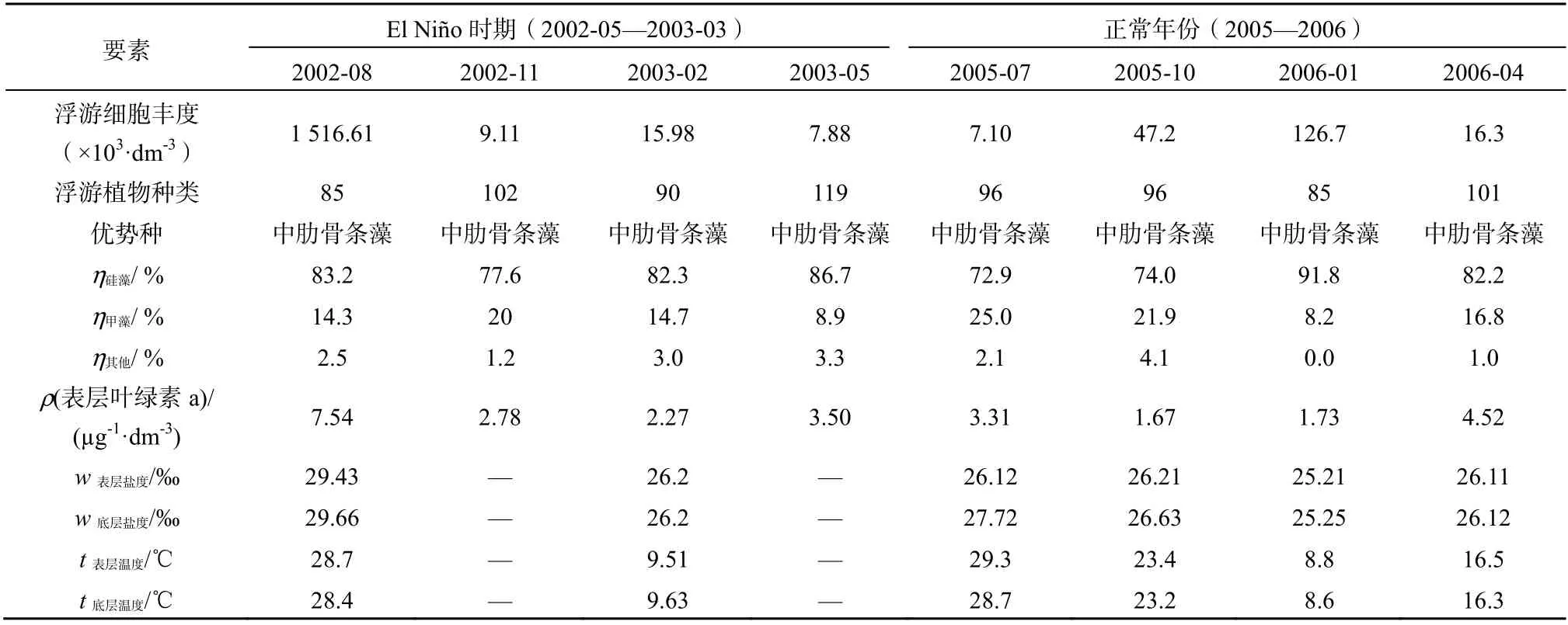
表3 三门湾EI Niño时期不同月份的浮游植物细胞丰度、种数和叶绿素a、初级生产力与正常年份对比 Table 3 Comparisons of cell abundance, species, chlorophyll a, primary productivity, temperature and salinity in the El Niño period with those in normal years
对比2002—2003年和2005—2006年,三门湾表层和底层温度盐度差异较大,海水温度和盐度是影响浮游植物生长繁殖的主要因子,不同类型的El Niño事件下底层盐度和温度场的特征不同,其温盐变化主要受浙江沿岸流和夏季台湾暖流的制约,明显具有外海水入侵的迹象[17]。分析了表3中2002—2003年与2005—2006年生物实测数据,优势物种以硅藻为主,浮游植物的分布特征主要受到硅藻的影响。在El Niño盛期,浮游植物细胞丰度剧增,而浮游植物群集多样性程度及物种均一性降低,中肋骨条藻类优势地位没有变化,甲藻在组成中的比例增加、物种多样性明显减少,这是中肋骨条藻类的绝对优势地位以及甲藻数量上升分布不均衡导致。一般来说,浮游植物的生长呈季节性变化(除了在赤道上升流区),三门湾浮游植物细胞丰度在不同年份的4个季节分布并不总是夏季高、冬季低,浮游植物种类数亦是这样,有明显的年度之间差异,历史资料也佐证了这一点。1982—1983年强El Niño现象(1982-09—1983-03),使当时赤道东太平洋水温比常年高出4 ℃,强El Niño现象持续近2年。1982年冬季12月三门湾网采集浮游植物细胞丰度(2.34×108(×103·dm-3))[19],为周年观察值最高,表明对于三门湾海域浮游植物生产力的年度间差异影响最大的是El Niño现象,El Niño的影响在某种程度上胜过年度间的季节温差。
3 结论
(1)应用三门湾沉积地层中长链烯酮不饱和指数(Uk37)与表层海水温度SST的关系重建了历史上的表层海水古温度,Uk37估算的温度清晰地捕捉了海湾温度SST变化记录与El Niño/La Niña有关年间变化;估算的古温度(t估)在15.19~17.61 ℃波动,并与2002年实测海水温度对比,发现Uk37估算温度与秋季海水实测温度接近,可能秋季是颗石藻群落旺发季节。
(2)3种生物标志物总含量(菜子甾醇+甲藻甾醇+长链烯酮)的变化范围为710.7~6998.5 ng·g-1,生物标志物估算的总初级生产力在近180年内总体呈上升趋势,单一浮游植物标志物估算的硅藻相对比例高,甲藻次之,硅藻/甲藻比值在2.1~7.2波动,表现出三门湾浮游植物对海洋环境变化的敏感性,El Niño/La Niña事件影响到浮游植物细胞丰度变化、不同浮游植物单体增减,特别是群落结构变化(硅藻与甲藻的关系)与El Niño/La Niña事件有密切关联。
(3)利用三门湾2个年份的实测水温、盐度和浮游植物相关数据来分析群落结构变化与厄尔尼诺/正常年份的相关程度。研究表明,在El Niño盛期,浮游植物细胞丰度剧增,浮游植物种类数及群集多样性程度及物种明显降低,中肋骨条藻类优势地位没有变化,这是中肋骨条藻类的绝对优势地位以及甲藻数量降低分布不均衡导致;研究还发现,三门湾浮游植物细胞丰度在不同年份的4个季节分布并不总是夏季高、冬季低,浮游植物种类数亦是这样,有明显的年际变化,三门湾历史资料也佐证了这一点(1982年强El Niño),以上反映了三门湾海域浮游植物生产力的年际和季节间的差异影响最大的是El Niño现象,El Niño的影响在某种程度上胜过年度间的季节温差。
[1] McPHADEN M J, ZHANG D X. Slowdown of the meridinal overturning circulation in the upper Pacific Ocean[J]. Nature, 2002, 415: 603-608.
[2] 颜尤明. 厄尔尼诺现象对闽南-台湾浅滩渔场灯光围网渔业影响的研究[J]. 福建水产, 2006,4: 11-15.
[3] ROPELEWSKI C F, HALPERT M S. Global and regional scale precipitation patterns associated with the El Niño/Southern Oscillation[J]. Monthly Weather Review, 1987, 115: 1606-1626.
[4] TURK D, McPHADEN M J, BUSALACCHI A J, et al. Remotely sensed biological production in the Equatorial Pacific[J]. Science, 2001, 293: 471-474.
[5] BEHRNFELD M J, O'MALLEY R T, SIEGEL D A, et al. Climate-driven trends in contemporary ocean productivity[J]. Nature, 2006, 444: 752-755.
[6] RODBELL D T, SELTZER G O, ANDERSON D M, et al. An 15000 year record of Ei Nio-driven alleviation in southwestern Ecuador[J]. Science, 1999, 283:516-520.
[7] RITTENOUR T M, BRIGHAM-GRETTE J, MANN M E. El Nio-Like climate teleconnections in new englang during the lake Pleistocene[J]. Science, 2000, 288: 1039-1042.
[8] SHULMEISTER J, LESS B G. Pollen vvidence from tropical Austrfor the onset of and ENSO-dominatrd climate at c. 4000 BP[J]. The Holocene, 1995, 5: 10-18.
[9] BIONDI F, GERSHUNOW A, CAYAN D R. North Pacific decadal climate variability since1661[J]. Climate, 2001, 14:5-10.
[10] VOLKMAN J K, BARRETT S M, BLACKBUM S I, et al. Microal galbiomarkers a review of recent research developments[J]. Organic Geochemistry, 1998, 29:1163-1179.
[11] ZIMMEMAN A R, CANUEL E A. Sediment geochemical records of eutrophication in the mesohaline Chesapeake bay[J]. Linnology and Oceanography, 2002, 47:1084-1093.
[12] EGLINTON T I, CONTE M H, EGLINTON G, et al. Proceedings of a workshop on alkenone-based paleoceanographic indicators[J]. Geochemistry Geophysics Geosystens, 2001, 2: 10. 1029/2000GC000122.
[13] ELEJERO C, GRIMALT J O. The correlation between the Uk37index and sea surface temperatures in the warm boundary: the South China Sea[J]. Geochim. Cosmochim. Acta, 1997, 22 (61): 4789-4797.
[14] EPSTEIN B L, HONDT S D, HARGRAVES P E. The possible metabolic role of C37alkenones in Emiliania huxleyi[J]. Organic Geochemistry, 2001: 32867-32875.
[15] STEINKE S, KIENAST M, PFLAUMANN F U, et al. A high-resolution sea-surface temperature record from the tropical South China Sea(16 500~3 000yr B.P.) [J]. Quaternary Research, 2001, 55: 352-362.
[16] ZHAO M X, EGLINTO G, READ G. An alkenone (Uk37) quasi-annual seaface temperature record (A.D.1440 to 1940) using varved sdiments from the Santa Barbara Basin[J]. Orgnic Geochemisry, 2000, 31: 903-917.
[17] 宁修仁. 乐清湾、三门湾养殖生态和养殖容量研究与评价[M]. 北京:海洋出版社, 2005: 111-168.
[18] 张冲,赵景波. 厄尔尼诺/拉尼娜事件对陕西气候的影响[J]. 陕西师范大学学报: 自然科学版,2010, 38:98-104.
[19] 浙江省海岸带和海涂资源综合调查报告编写委员会. 浙江省海岸带和海涂资源综合调查报告[M]. 北京:海洋出版社,1985.
[20] 陈荣华,赵庆英,王炳泉,等. 浙江三门湾柱样沉积中长链烯酮、正构烷烃和多环芳烃组成、分布特征及对厄尔尼诺/拉尼娜事件的追踪[J]. 生态环境学报. 2012, 5:890-896.
