湘西北河谷特殊生境植物多样性和区系成分及其与土壤环境因子的相关性
2013-07-10陈功锡张代贵
徐 亮,陈功锡,①,张 洁,邓 涛,张代贵
(1. 吉首大学植物资源保护与利用湖南省高校重点实验室,湖南吉首416000;2. 中国科学院昆明植物研究所生物多样性与生物地理学重点实验室,云南昆明650201)
特殊生境即特殊生态环境的简称,是指那些在结构与功能上具有明显的特殊性(或异质性),并导致生态元的数量或品质明显不同的生态环境。特殊生境极易孕育特有现象[1],它可通过与周围生境的突然间断性产生作用,如地理地形条件形成的峡谷(悬崖)、山顶(孤山)和特殊的土壤条件(如岩层露面和碎石夹杂的土壤等)[2-3]。河谷是山地区域江流深切形成的一个自然区域,属典型的特殊生境。自20 世纪80年代以来,国内学者对干热河谷特殊生境植物区系特征、性质和起源进行了研究[4-8],认为深陷的河谷地貌和特殊的气候条件是形成其独特植物区系的原因之一;河谷特殊生境形成的植物区系对整个区域植物多样性和生态功能都具有举足轻重的作用。
有关植物与环境因子间相互关系的讨论较为广泛[9-12],环境对于植物的生长、繁殖和分布都有重要作用,被视为气候、生物有机体、地形、母质和时间的一个函数[13]。近年来,涉及土壤、生境特点与植物分布之间关系的研究不少,如对长白山北坡苔藓、草本和木本植物分布与环境关系的研究[14-16],但有关植物区系成分与环境间关系的研究尚不多见。
猛洞河与德夯是湘西北河谷特殊生境的典型代表,是研究植物区系成分与环境间关系的较理想场所。陈功锡等[17]认为:德夯河谷的生境异质性能在一定程度上影响区系成分的变化,但两者之间的具体关系还不清楚。为此,作者拟对猛洞河与德夯河谷不同小生境内的植物种类进行全面调查,同时对各小生境的土壤因子进行测定,通过冗余分析(redundancy analysis,RDA)探讨河谷特殊生境与植物多样性、植物区系成分之间的关系,进一步加深对河谷生境异质性与植物组成和分布的认识,为湘西北河谷特殊生境的植物多样性形成与维持理论研究奠定基础。
1 研究区概况和研究方法
1.1 研究区概况
猛洞河地处湖南省武陵山腹地,地理坐标为北纬28°36' ~28°56'、东经 109°14' ~109°57',地跨永顺县和古丈县;属中亚热带温润季风气候区,岩溶作用强盛,为中低山溶丘洼地峡谷地貌;境内溶洞密布,两岸石壁耸峙狭陡呈“U”形河谷,落差达100 ~200 m,海拔350 ~650 m;年均温约16 ℃,年降水量1 300 ~1 600 mm;植被类型以亚热带常绿阔叶林为主。
德夯河谷位于湖南省吉首市境内,地理坐标为北纬 28°15' ~28°43'、东经 109°30' ~109°45';地貌为典型的喀斯特岩溶峡谷景观,境内坡陡壁峭,呈半封闭式的“V”形河谷,海拔250 ~790 m;年均温16 ℃ ~17 ℃,年降水量1 200 ~1 600 mm;植被类型主要为中亚热带常绿阔叶林、暖性针叶林及竹林[18]。
1.2 研究方法
1.2.1 生境划分及植物种类调查 在2 个河谷中划分出凹槽、干旱石壁、滴水岩壁、石土坡、瀑布、微土台、溪沟、岩溶洞穴和河水浸没带 9 个小生境类型[18];在每一小生境中植被保存较好、生境特征明显的地段均划定10 个调查样地,样地面积视具体情况而定,一般乔木样地面积为10 m×10 m,灌木及草本样地的面积分别为5 m×5 m 和1 m×1 m。石土坡生境面积较大,采用样方和样带相结合的方法进行调查;其他小生境则进行全面调查,范围为小生境主体部分及左右(或上下)辐射1 m 的区域,记录物种名称、数量、高度等指标[19]。对重要植物则采集标本,凭证标本存于吉首大学生物系植物标本室(JIU)。
1.2.2 土样采集及土壤指标测定 受地形和小气候影响,各小生境的土层均较薄,植被地带性不明显,构成以灌木和草本为优势的非地带性群落。基于此种情况,在进行土壤取样时直接从各小生境样地优势植物的根部挖取表层土壤(土壤深度5 ~10 cm)100 ~200 g,每个小生境确定5 个样地,每个样地各取上、中、下3 个样点的土壤,除去其中的砂石及植物残体并混合均匀后带回实验室进行相关指标分析。
采用烘干法[20]测定土壤含水量;使用酸度计测定土壤 pH 值[21];先用 MDS-6 型微波消解仪(上海新仪微波化学科技有限公司)对土样进行消解,然后用iCAP 6000 SERIES 电感耦合等离子体原子发射光谱仪(英国热电元素公司)测定各矿质元素含量。各指标均重复测定3 次。
1.3 数据处理和分析
采用Excel 2003 软件对调查数据进行统计分析,采用Canoco (Version 4.0)分析软件对土壤因子和物种数量的相关性进行 RDA 分析和排序,使用CanoDraw (Version 4.0)软件绘制 RDA 排序图。
2 结果和分析
2.1 土壤组成特征
湘西北猛洞河与德夯河谷9 个小生境土壤含水量、pH 值及矿质元素含量的测定结果见表1。
2.1.1 土壤含水量分析 由表1 可知:9 个小生境的土壤含水量差异较大。其中,瀑布生境的土壤含水量最高,达51.9%,这与瀑布水流充沛、湿度较大有关;溪沟和河水浸没带的土壤含水量分别为48. 2% 和45.4%,由于溪沟和河水浸没带常有季节性积水,相当于河谷的湿地,故含水量较高;滴水岩壁土壤含水量为43.4%,此生境常年滴水,但风化程度较低,土层稀薄,储水能力较差;岩溶洞穴土壤含水量为30.4%,由于地下和地表水对岩石进行溶蚀,从而产生特殊的地质地貌,在其晚期形成过程中完全脱离地下水位而导致土壤含水量降低;凹槽、微土台和石土坡土壤含水量较低,分别为 27.3%、26.7%和 26.6%;9 个小生境中干旱石壁土壤含水量最低,仅为13.0%,因该生境受阳光直射时间最长,且温度最高、土壤稀薄、储水能力很差,导致土壤含水量极低。
表1 湘西北猛洞河与德夯河谷不同小生境土壤含水量、pH 值及矿质元素含量的测定结果(±SD)Table 1 Determination result of water content,pH value and mineral element content in soil of different micro-habitats in valley of Mengdonghe and Dehang of Northwestern Hu’nan Province (±SD)
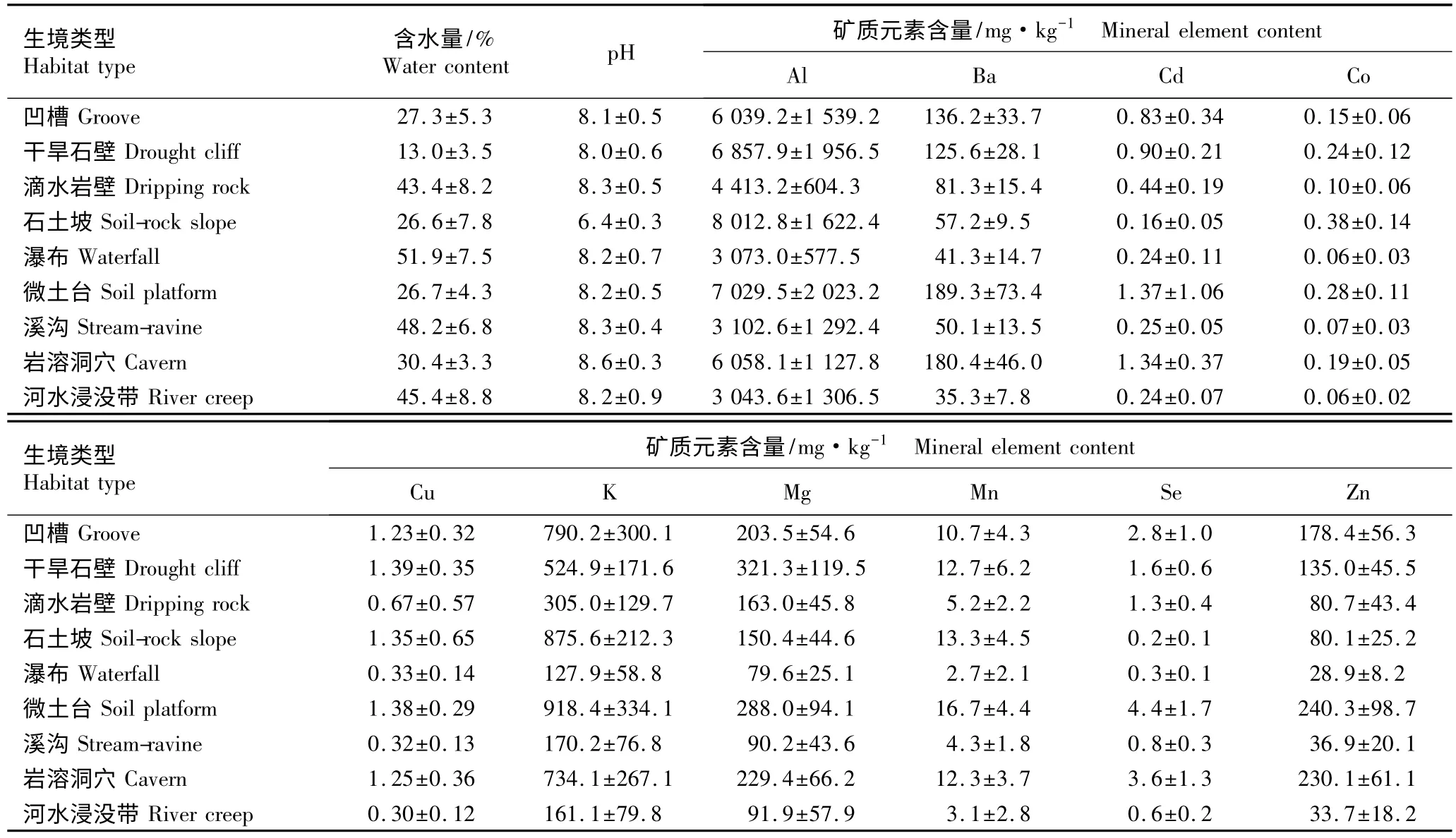
表1 湘西北猛洞河与德夯河谷不同小生境土壤含水量、pH 值及矿质元素含量的测定结果(±SD)Table 1 Determination result of water content,pH value and mineral element content in soil of different micro-habitats in valley of Mengdonghe and Dehang of Northwestern Hu’nan Province (±SD)
生境类型Habitat type 含水量/%Water content pH 矿质元素含量/mg·kg-1 Mineral element content Al Ba Cd Co凹槽 Groove 27.3±5.3 8.1±0.5 6 039.2±1 539.2 136.2±33.7 0.83±0.34 0.15±0.06干旱石壁 Drought cliff 13.0±3.5 8.0±0.6 6 857.9±1 956.5 125.6±28.1 0.90±0.21 0.24±0.12滴水岩壁 Dripping rock 43.4±8.2 8.3±0.5 4 413.2±604.3 81.3±15.4 0.44±0.19 0.10±0.06石土坡 Soil-rock slope 26.6±7.8 6.4±0.3 8 012.8±1 622.4 57.2±9.5 0.16±0.05 0.38±0.14瀑布 Waterfall 51.9±7.5 8.2±0.7 3 073.0±577.5 41.3±14.7 0.24±0.11 0.06±0.03微土台 Soil platform 26.7±4.3 8.2±0.5 7 029.5±2 023.2 189.3±73.4 1.37±1.06 0.28±0.11溪沟 Stream-ravine 48.2±6.8 8.3±0.4 3 102.6±1 292.4 50.1±13.5 0.25±0.05 0.07±0.03岩溶洞穴 Cavern 30.4±3.3 8.6±0.3 6 058.1±1 127.8 180.4±46.0 1.34±0.37 0.19±0.05河水浸没带 River creep 45.4±8.8 8.2±0.9 3 043.6±1 306.5 35.3±7.8 0.24±0.07 0.06±0.02生境类型Habitat type矿质元素含量/mg·kg-1 Mineral element content Cu K Mg Mn Se Zn凹槽 Groove 1.23±0.32 790.2±300.1 203.5±54.6 10.7±4.3 2.8±1.0 178.4±56.3干旱石壁 Drought cliff 1.39±0.35 524.9±171.6 321.3±119.5 12.7±6.2 1.6±0.6 135.0±45.5滴水岩壁 Dripping rock 0.67±0.57 305.0±129.7 163.0±45.8 5.2±2.2 1.3±0.4 80.7±43.4石土坡 Soil-rock slope 1.35±0.65 875.6±212.3 150.4±44.6 13.3±4.5 0.2±0.1 80.1±25.2瀑布 Waterfall 0.33±0.14 127.9±58.8 79.6±25.1 2.7±2.1 0.3±0.1 28.9±8.2微土台 Soil platform 1.38±0.29 918.4±334.1 288.0±94.1 16.7±4.4 4.4±1.7 240.3±98.7溪沟 Stream-ravine 0.32±0.13 170.2±76.8 90.2±43.6 4.3±1.8 0.8±0.3 36.9±20.1岩溶洞穴 Cavern 1.25±0.36 734.1±267.1 229.4±66.2 12.3±3.7 3.6±1.3 230.1±61.1河水浸没带 River creep 0.30±0.12 161.1±79.8 91.9±57.9 3.1±2.8 0.6±0.2 33.7±18.2
2.1.2 土壤pH 值分析 由表1 可见:猛洞河和德夯河谷9 个小生境的土壤大部分偏碱性。其中,岩溶洞穴的土壤pH 值最高,为pH 8.6;石土坡的土壤pH 值最低,仅为pH 6.4。河谷土壤是石灰岩母质与其他自然因素相互作用形成的,其土壤大部分偏碱性。然而阴湿山坡的土壤是经过淋溶作用形成的,在淋溶过程中由于雨水吸收空气CO2后渗入土壤并与土壤有机质或矿物质分解所产生的各种酸相混合,使其酸性更强,可将土壤内的石灰质和其他可溶性盐溶解,导致石土坡生境的土壤偏酸性。
2.1.3 土壤矿质元素含量分析 由表1 可见:9 个生境土壤中 Al、Ba、Cd、Co、Cu、K、Mg、Mn、Se 和 Zn 的含量有明显差异。石土坡生境土壤中Al 含量最高、Cd含量最低;其他8 个小生境的土壤中均为Al 含量最高、Co 含量最低;Al 和Ba 含量在石土坡土壤中最高、在河水浸没带最低;Cd 含量在微土台土壤中最高、在河水浸没带和瀑布中最低;Co 含量在石土坡土壤中最高、在河水浸没带和瀑布中最低;Cu 含量在干旱石壁土壤中最高、在河水浸没带中最低;K、Mn 和Zn 含量在微土台土壤中最高、在瀑布中最低;Mg 含量在干旱石壁土壤中最高、在瀑布中最低;Se 含量在微土台土壤中最高、在石土坡中最低。
2.2 物种多样性分析
2.2.1 物种多样性特征分析 湘西北猛洞河与德夯河谷9 个小生境中植物物种组成的分析结果见表2。
由表2 可知:各生境中植物物种数量有明显差异。其中,石土坡生境中植物种类数最多,达735 种,隶属于132 科397 属,占河谷植物总种数的62.8%;该生境土层较厚、腐殖质含量较高、含水量适中,能为各种植物提供适宜的生存条件;分布有以湘西青冈(Cyclobalanopsis xiangxiensis C. J. Qi et Q. Z. Lin)和湘西石栎(Lithocarpus hunanensis C. J. Qi)等特有种为建群种的群落,是河谷植物分布的主要载体。

表2 湘西北猛洞河与德夯河谷不同小生境植物科、属和种的数量Table 2 Number of family,genus and species of plant in different micro-habitats in valley of Mengdonghe and Dehang of Northwestern Hu’nan Province
河水浸没带分布的植物种类数位居第2,有242种,隶属于79 科197 属,占总种数的20.68%;该生境处于较为开阔地带,受河水高涨浸没和生境破坏的影响其植被以次生性为主,同时大量河谷外种类通过河水及地形的作用渗入并定居于该小生境中,导致该小生境中物种多样性相对较高;常见的种类有苍耳(Xanthium sibiricum Patrin ex Widder)、车前(Plantago asiatica Linn.)和酢浆草(Oxalis corniculata Linn.)等。
微土台生境中分布有216 种植物,隶属于55 科159 属,占总种数的 18. 46%;代表种类有雾水葛〔Pouzolzia zeylanica (Linn.) Benn.〕和 点 地 梅〔Androsace umbellata (Lour.)Merr.〕等。
溪沟生境中分布有168 种植物,隶属于59 科125属,占总种数的14. 36%;代表种类有光叶槭(Acer laevigatum Wall.)和块节凤仙花(Impatiens pinfanensis Hook. f.)等。
干旱石壁生境中分布有144 种植物,隶属于64科117 属,占总种数的12.3%;由于该生境土壤含水量很低,保水功能也较差,一般植物很难生存,只有青檀 (Pteroceltis tatarinowii Maxim.)、吉首紫菀 (Aster jishouensis W. P. Li et S. X. Liu)和柄果薹草(Carex stipitinux C. B. Clarke ex Franch.)等种类的分布。
凹槽小生境中分布有80 种植物,隶属于43 科69属,占总种数的6.84%;该生境中常见的种类有密脉木(Myrioneuron faberi Hemsl.)和动蕊花〔Kinostemon ornatum (Hemsl.)Kudô〕等。
瀑布生境中分布有49 种植物,隶属于30 科41属,占总种数的4.19%;因该生境中具有高浓度的离子、高冲击力的水流和碳酸钙沉积,成为多数高等植物难以生存的一类特殊生境;仅分布有黄花狸藻(Utricularia aurea Lour.)和滴水珠(Pinellia cordata N.E. Brown)等种类。
滴水岩壁生境中分布有24 种植物,隶属于18 科22 属,占总种数的2.05%;该生境土壤含水量高且土层薄,不适宜乔木及灌木种类的生长,仅分布一些草本植物,如三脉紫菀(Aster ageratoides Turcz.)、永顺楼梯草(Elatostema yungshunense W. T. Wang)和 1 种过路黄属植物(Lysimachia sp.)等。
岩溶洞穴生境中分布的植物种类最少,仅有6种,隶属于6 科6 属,仅占总种数的0.51%;由于洞穴环境中温度较恒定、湿度相对较大且光照强度较小,大多数植物不适应此种生境;但有些植物种类对洞穴环境特别适应,甚至仅见于洞穴中且离开洞穴环境不能正常生长[22],如苦苣苔科(Gesneriaceae)的柔毛半蒴苣苔(Hemiboea mollifolia W. T. Wang)等。
2.2.2 物种适应性范围分析 物种的分布辐射性(diathermaneity)是指在一定区域内某个物种分布的小生境数量,为植物分布适应性的重要指标之一;辐射性越大物种适应性越强,反之则越弱;仅辐射1 个小生境的物种可以初步定为这一区域该生境的生境专有种。统计结果显示:在9个小生境中,分布辐射1 个小生境(石土坡)的种类有724 种,占总种数的61.8%;分布辐射2 个小生境(石土坡和微土台)的种类有305 种,占总种数的26.0%;分布辐射1 个和2个小生境的种类共有1 029 种,占总种数的87.8%。仅有蝴蝶花(Iris japonica Thunb.)1 种的适应范围较大,分布辐射达7 个小生境,仅在干旱石壁和岩溶洞穴生境中没有分布。
2.3 植物区系成分与小生境的关系
2.3.1 植物区系成分分析 根据吴征镒[23]和李锡文[24]对植物科属的分布区类型划分原则,对湘西北猛洞河与德夯河谷9 个小生境中植物科、属的地理分布区类型进行分析,结果见表3。
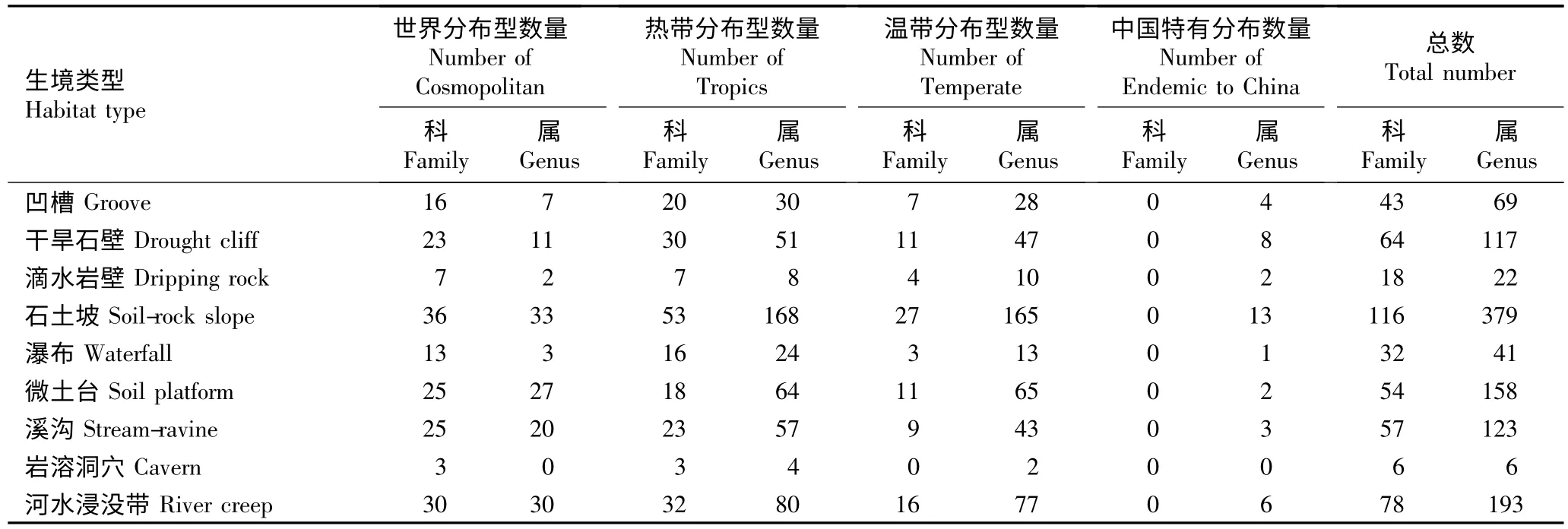
表3 湘西北猛洞河与德夯河谷不同小生境植物科、属的分布区类型统计Table 3 Statistics of areal-type of family and genus of plants in different micro-habitats in valley of Mengdonghe and Dehang of Northwestern Hu’nan Province
由表3 可知:在科级水平上,河水浸没带生境中分布着较多的世界分布型科,这可能是由于大量河谷外植物的繁殖体利用水体进行传播,退洪后便固着(营养体)或残留(种子)在河水浸没带上,在适宜条件下生长并定居与此。各小生境中热带成分科的数量均明显多于温带成分科,表现出较强的热带性质,这可能是由于河谷是一个相对稳定的生境,水热保持能力较强且常年处于温热潮湿的环境下,长期的适应进化导致其中的热带分布物种逐渐增多;典型的有樟科(Lauraceae)的润楠叶木姜子(Litsea machiloides Yen C. Yang et P. H. Huang)、壳斗科(Fagaceae)的湘西青冈和桑科(Moraceae)的岩木瓜(Ficus tsiangii Merr.ex Corner)等热带分布种类。
在属级水平上,除滴水岩壁和微土台生境中温带成分型属的数量稍高于热带成分型属外,其他小生境中热带成分型属的数量均略高于温带成分型属,而滴水岩壁和微土台小生境中温带成分进一步增加。在石土坡、凹槽和干旱石壁等小生境中一些典型的温带分布型属最为常见,如杜鹃花属(Rhododendron Linn.)、百合属(Lilium Linn.)和胡颓子属(Elaeagnus Linn.)等;在溪沟、瀑布和河水浸没带小生境中可以见到典型的北温带分布型属,如枸杞属(Lycium Linn.)、荚蒾属(Viburnum Linn.)、山茱萸属(Cornus Linn.)、蔷薇属(Rosa Linn.)、鸢尾属(Iris Linn.)和紫菀属(Aster Linn.)等。在滴水岩壁和微土台小生境中常见有接骨木属(Sambucus Linn.)、婆婆纳属(Veronica Linn.)、马桑属(Coriaria Linn.)、樟属(Cinnamomum Schaeff.)和兔儿风属(Ainsliaea DC.)等属的种类,故这2 类小生境的温带区系成分相对明显。
2.3.2 植物区系成分与小生境土壤环境因子的RDA排序 湘西北猛洞河与德夯河谷9 个小生境的植物区系成分与土壤环境因子的RDA 分析结果见表4 和表5;植物科和属的分布区类型数量与各生境中土壤环境因子的RDA 排序图见图1 和图2。

表4 湘西北猛洞河与德夯河谷不同小生境植物科、属的分布区类型与土壤环境因子的RDA 分析结果Table 4 RDA analysis result of areal-type of plant family and genus with soil environmental factors in different micro-habitats in valley of Mengdonghe and Dehang of Northwestern Hu’nan Province

表5 湘西北猛洞河与德夯河谷不同小生境植物科、属分布区类型的RDA 排序轴与土壤环境因子的相关系数Table 5 Correlation coefficient of RDA ordination axes of areal-type of plant family and genus with soil environmental factors in different micro-habitats in valley of Mengdonghe and Dehang of Northwestern Hu’nan Province
由表4 可见:在科级水平上,区系成分信息总变异中的75.9%能够被土壤中Ba 含量、土壤pH 值和土壤含水量等12 个变量解释;其中第1 排序轴解释了49.9%的变异信息,第2 排序轴解释了约26.0%的变异信息。在属级水平上,区系成分信息总变异中的88.9%能够被土壤的12 个变量解释;其中,第1 排序轴解释了75.7%的变异信息,第2 排序轴解释了约13.2%的变异信息。
由表5 可见:土壤 Cd、Se 和 Zn 含量在科、属水平上均与第1 轴呈显著正相关,Ba 含量在科、属水平上均与第2 轴呈显著正相关,土壤pH 值在科级水平上与第2 轴呈显著负相关。表明在12 个土壤环境因子中,土壤 Cd、Se、Zn 和 Ba 含量以及土壤 pH 值是影响湘西北河谷植物区系组成的主要土壤因子。
从科的RDA 排序图(图1)可见:在9 个小生境中,东亚北美间断分布型、热带亚洲至热带大洋洲分布型、北温带分布型、热带亚洲分布型、热带亚洲至热带非洲分布型、东亚分布型和旧世界温带分布型科的数量与土壤Ba 含量呈正相关,而与土壤Zn、Cd 和Se含量均呈负相关;热带亚洲至热带美洲间断分布型和旧世界温带分布型科的数量与土壤Zn、Cd 和Se 含量均呈负相关,与土壤pH 值呈正相关;泛热带分布型和世界分布型科的数量与土壤pH 值以及Zn、Cd 和Se含量均呈正相关,与土壤Ba 含量呈负相关。
从属的RDA 排序图(图2)可见:在9 个小生境中,中国特有分布型、温带亚洲分布型、热带亚洲至热带大洋洲分布型、东亚北美间断分布型、东亚分布型、地中海至中西亚分布型、旧世界热带分布型、世界分布型和旧世界温带分布型属的数量均与土壤Ba 含量呈正相关,与土壤Cd、Se 和Zn 含量呈负相关;北温带分布型、泛热带分布型和热带亚洲分布型属的数量与土壤Cd、Se 和Zn 含量呈正相关,与土壤Ba 含量呈负相关;热带亚洲至热带美洲间断分布型和热带亚洲至热带非洲分布型属的数量与土壤Ba、Cd、Se 和Zn 含量均呈负相关。

图1 湘西北猛洞河与德夯河谷不同小生境植物科的分布区类型数量与土壤环境因子的RDA 排序图Fig. 1 RDA ordination diagram of plant family areal-type number with soil environmental factors in different micro-habitats in valley of Mengdonghe and Dehang of Northwestern Hu’nan Province
以上结果说明:在科级和属级水平上,生境异质性均能在一定程度上影响植物区系成分的变化。
3 讨论和结论
3.1 生境异质性与物种组成的关系
影响物种多样性的因素很多,如水热条件、土壤深浅和肥力状况等都直接影响植物的栖息定居,而异质的环境因子是大量物种得以在群落中稳定共存的重要因素[25]。异质性程度越高,意味着小生境及小气候条件越丰富,能为具有不同生活史策略和生理生态要求的物种提供定居的生态位,有利于生物多样性的维持[26-27]。俞筱押等[28]和赵存玉等[29]分别调查了云南石林和科尔沁沙地草场的小生境植物,认为不同的小生境类型对植物多样性的影响非常明显,本研究结果也支持这一结论。从本研究结果来看,湘西北猛洞河与德夯河谷不同小生境的植物多样性差异较大,其中石土坡的物种多样性水平最高(共分布有735 种植物),占河谷中植物总种数的62.8%,是河谷植物的主要载体;该生境土层较厚、腐殖质含量较高、含水量适中,能为各种植物提供适宜的生存条件。岩溶洞穴生境中物种多样性水平最低(仅6 种),仅占河谷植物总种数的0.5%。从分布辐射性可见:分布辐射1 个小生境的种类有724 种,占总种数的61.8%,比例相对较高,说明在河谷这一特殊生境中物种分布较专一、抗干扰能力较差,不利于种群和物种多样性的长期维持。

图2 湘西北猛洞河与德夯河谷不同小生境植物属的分布区类型数量与土壤环境因子的RDA 排序图Fig. 2 RDA ordination diagram of plant genus areal-type number with soil environmental factors in different micro-habitats in valley of Mengdonghe and Dehang of Northwestern Hu’nan Province
3.2 生境异质性与植物区系成分的关系
目前,人们所揭示的植物区系的分布规律主要决定于大气候,亦即呈现一定的地带性规律。但在长期的进化过程中,植物为适应各种复杂的生境变化也可能出现植物区系的分化或特化[30-31];也有学者认为:植物区系成分在形态-地理学、细胞-地理学、分子-(生物)地理学等水平上均存在地带性分化特点[32]。作者的长期观察与研究结果显示:除主要的地带性区系成分以外,在湘西北尤其是在河谷特殊生境中分布着丰富且多样化的非地带性植物区系成分[17],它们的形成可能受小气候的影响,它们的存在也在一定程度上决定了该区域植物区系的特征与性质。
沈泽昊等[33]利用主成分分析方法(PCA)研究了中国亚热带区域植物区系地理成分构成的相关性和空间分异特征,结果表明:经PCA 排序后,属的分布区类型可分为热带分布、地中海-中亚分布、温带分布和东亚分布4 个类型;同时,纬度的梯度变化对区系地理成分分布有突出影响,表现出与纬度的极显著负相关性。由此可见,环境因子的改变会直接导致植物区系成分发生变化。
有“约束下的排序方法”之称的RDA 分析法能直接将环境变量作为约束条件考虑到排序分析中,从而大幅度消减环境变量的规模[34]。本研究结果表明:生境中不同的土壤环境因子对植物区系的组成有显著影响,且在科、属2 级水平上的影响程度有差异。在科级水平上,区系成分信息总变异中的75.9%能够被所选的12 个土壤环境因子变量解释,其中特征值大于0.1 的有5 个变量,分别是土壤pH 值以及土壤Ba、Zn、Cd 和 Se 含量;在属级水平上,区系成分信息总变异中的88.9%能够被12 个土壤环境因子变量解释,其中特征值大于0.1 的有4 个变量,分别是土壤Ba、Zn、Cd 和Se 含量。由此也可看出:物种在进化过程中具有生态位保守性(niche conservatism),且在属级水平上物种生态位的保守性高于科级水平。因此,趋同进化也应该是湘西北植物区系形成的一个重要原因,而生境异质性在一定程度上影响该区域植物区系成分的变化,但这种影响的普遍性及其作用机制与途径还有待进一步研究探索。
致谢:在本文写作过程中得到吉首大学生物资源与环境科学学院何兴兵副教授的大力支持,谨此致谢!
[1]张宏达. 植物的特有现象与生物多样性[J]. 生态科学,1997,16(2):9-17.
[2]BASKIN J M,BASKIN C C. Endemism in rock outcrop plant communities of unglaciated eastern United States:an evaluation of the roles of the edaphic,genetic and light factors[J]. Journal of Biogeography,1988,15:829-840.
[3]GILBERT E. Flora of the West Fork of Oak Creek,Coconino County,Arizona[M]. Tempe:Arizona State University,2004:1-119.
[4]周立江,管中天. 金沙江河谷苏铁天然植物群落的研究[J]. 云南植物研究,1985,7(2):153-168.
[5]欧晓昆. 元谋干热河谷植物区系研究[J]. 云南植物研究,1988,10(1):11-18.
[6]曹 敏,金振洲. 云南巧家金沙江干热河谷的植被分类[J]. 云南植物研究,1989,11(3):324-336.
[7]曹永恒,金振洲. 云南潞江坝怒江干热河谷植被研究[J]. 广西植物,1993,13(2):132-138.
[8]金振洲,杨永平,陶国达. 华西南干热河谷种子植物区系的特征、性质和起源[J]. 云南植物研究,1995,17(2):129-143.
[9]UKPONG I E. Salinity in the Calabarmangrove swamp,Nigeria[J].Mangroves and Salt Marshes,1997,1:211-218.
[10]ABD EL-GHANI M M,AMER W M. Soil vegetation relationships in a coastal desert plain of southern Sinai,Egypt[J]. Journal of Arid Environments,2003,55(4):607-628.
[11]EL-DEMERDASH M A,HEGAZY A K,ZILAY A M. Vegetationsoil relationships in Tihamah coastal plains of Jazan region,Saudi Arabia[J]. Journal of Arid Environments,1995,30(2):161-174.
[12]CRITCHLEY C N R,CHAMBERS B J,FOWBERT J A,et al.Plant species richness,functional type and soil properties of grasslands and allied vegetation in English environmentally sensitive areas[J]. Grass and Forage Science,2002,57:82-92.
[13]JAFARIA M,ZARE CHAHOUKI M A,TAVILI A,et al. Effective environmental factors in the distribution of vegetation types in Poshtkouh rangelands of Yazd Province (Iran)[J]. Journal of Arid Environments,2004,56(4):627-641.
[14]GUO S L,CAO T. Distribution patterns of ground moss species and its relationships with environmental factors in Changbai Mountain.Northeast China[J]. Journal of Integrative Plant Biology,2001,43(6):631-643.
[15]郝占庆,郭水良. 长白山北坡草本植物分布与环境关系的典范对应分析[J]. 生态学报,2003,23(10):2000-2008.
[16]郝占庆,郭水良,叶 吉. 长白山北坡木本植物分布与环境关系的典范对应分析[J]. 植物生态学报,2003,27(6):733-741.
[17]陈功锡,邓 涛,张代贵,等. 湖南德夯风景区峡谷特殊生境植物区系与生态适应性初探[J]. 西北植物学报,2009,29(7):1470-1478.
[18]邓 涛. 河谷特殊生境物种多样性特征与生态适应性——以湘西北主要河谷为例[D]. 吉首:吉首大学生物资源与环境科学学院,2010:10-16.
[19]宋永昌. 植被生态学[M]. 上海:华东师范大学出版社,2001.
[20]华 孟,王 坚. 土壤物理学[M]. 北京:北京农业大学出版社,1993.
[21]蒋 实,徐争启,张江成. 四川省万源市土壤pH 值测定及土壤酸碱度分析[J]. 安徽农业科学,2009,37(25):12105-12108.
[22]许为斌. 广西溶岩洞穴植物初步研究[D]. 桂林:广西师范大学生命科学学院,2004.
[23]吴征镒. 中国种子植物属的分布区类型[J]. 云南植物研究,1991,13(增刊Ⅳ):1-139.
[24]李锡文. 中国种子植物区系统计分析[J]. 云南植物研究,1996,18(4):363-384.
[25]沈泽昊,张新时.三峡大老岭地区森林植被的空间格局分析及其地形解释[J]. 植物学报,2000,42(10):1089-1095.
[26]EHRENFELD J G. Microtopography and vegetation in Atlantic white cedar swamps:the effects of natural disturbances[J].Canadian Journal of Botany,1995,73:474-484.
[27]NAKAMURA E,YAJIMA T,KIKUCHI S. Structure and composition of riparian forests with special reference to geomorphic site conditions along the Tokachi river,northern Japan[J]. Plant Ecology,1997,133:209-219.
[28]俞筱押,李玉辉,马遵平. 云南石林喀斯特小生境木本植物多样性特征[J]. 山地学报,2007,25(4):438-447.
[29]赵存玉,王 涛,董治宝,等. 科尔沁沙地草场物种多样性特征及其与生境的关系[J]. 草业科学,2007,24(10):11-18.
[30]吴征镒,朱彦成. 云南植被[M]. 北京:科学出版社,1987.
[31]曾昭璇. 植被的地形分化论:以华南植被为例[J]. 华南师范大学学报:自然科学版,1981(2):88-102.
[32]崔大方,廖文波,王伯荪. 植物区系特征成分及地带性分化问题的探讨[J]. 中山大学学报:自然科学版,2000,29(5):73-77.
[33]沈泽昊,张新时. 中国亚热带地区植物区系地理成分及其空间格局的数量分析[J]. 植物分类学报,2000,38(4):366-380.
[34]PIERRE L. Study beta diversity:ecological variation partitioning by multiple regression and cannoical analysis[J]. Journal of Plant Ecology,2007,31(5):976-981.
猜你喜欢
——以贵阳花溪公园为例
