泽兰实蝇内生殖器官的结构及其发育状况
2013-07-05汪金蓉李正跃吴国星
高 熹,马 沙,汪金蓉,叶 敏,李正跃,吴国星
云南农业大学植物保护学院,云南昆明650201
泽兰实蝇Procecidochares utilisStone属双翅目实蝇科,是紫茎泽兰Eupatorium adenophorumSpreng.重要的专性寄生性天敌。美国、澳大利亚、新西兰和印度等国家曾先后利用该虫控制紫茎泽兰的危害(Dodd,1961;Rahman & Agarwal,1991)。我国于1894年首次从西藏聂拉木县将泽兰实蝇引进云南部分地区用于控制紫茎泽兰,现已在云南省约5000 km2范围内定殖,后被引进四川、贵州等地并形成自然种群(陈升碧和关德盛,1991;魏艺等,1989;周川江等,2003)。研究发现,泽兰实蝇通过幼虫蛀入紫茎泽兰幼嫩茎枝端部,蛀食并刺激被害部位膨大形成虫瘿,从而抑制紫茎泽兰种子的萌发和植株的分蘖,抑制其光合作用使节间缩短甚至枯死等,显著降低种子产量和萌发率以及分枝数(陈旭东和何大愚,1990;何大愚和梁家社,1988;刘文耀等,1991;张智英等,1988;郑征,1989;Bennett& Vanstaden,1986;Bess&Haramoto,1958)。泽兰实蝇对入侵生物紫茎泽兰的专性寄生可有效控制紫茎泽兰的扩散,是一种有效的生物防治手段。因此,研究泽兰实蝇的生物学及生理功能,可为提高泽兰实蝇的人工繁殖效率及生物防治效果奠定理论基础。
昆虫的生殖系统由外生殖器和内生殖器2个部分组成。外生殖器一般由腹部末端的几个体节和附肢组成,如产卵器、阴茎及抱握器等,用于完成雌雄虫的交配和受精作用;内生殖器主要是由中胚层形成的生殖腺和附腺,如卵巢、精巢、输卵管及输精管等,外胚层部分内陷形成管道,如中输卵管、阴道及射精管等,其作用是产生成熟的精子和卵细胞(王晓芳,2010),对内生殖器官结构及发育规律的了解是研究生殖生理的首要条件。昆虫种类不同,其生殖系统的差异很大(詹光杰,2009)。昆虫的生殖能力与其生殖器官的发育密切相关(Davey,1985)。国内外对泽兰实蝇的研究多集中于生物学特性和生防应用方面(马沙等,2012),尚未见有关其生殖系统发育的报道。鉴于此,本试验采用光学显微的方法对泽兰实蝇内生殖器官的结构和发育进行初步研究。
1 材料与方法
1.1 供试虫源
泽兰实蝇采自云南昆明呈贡县带有虫瘿的紫茎泽兰枝条,摘除部分枝叶后把虫瘿放入养虫笼(60 cm×60 cm×60 cm)中,让其自然羽化,收集羽化的泽兰实蝇作为供试虫源。
1.2 泽兰实蝇的饲养
泽兰实蝇羽化后,将雌雄虫分开饲养,在20℃、RH 75%、光周期L∶D=14∶10的条件下进行饲养。不提供寄主产卵,仅喂食10%蜂蜜水。
1.3 生殖系统的解剖与测量
自成虫羽化当天开始,雌雄成虫每天各取30头,解剖其生殖系统直至成虫正常死亡。用微型解剖针,将泽兰实蝇的头和胸除去,撕开腹部,取出生殖系统,用0.9%生理盐水清洗生殖系统,剔除附着在生殖系统的脂肪体和微气管。将解剖好的生殖器官移到立体解剖镜(Olympus Stemi 2000-C)下,对生殖系统中的卵巢、雌性附腺、受精囊、精巢、雄性附腺等进行拍照,并用附带的软件分析系统进行测量。长度为器官或腺体的基部到端部的距离;宽度是对器官的最宽处进行测量。
1.4 数据处理
利用DPS软件(唐启义和冯明光,2002)对不同发育日龄的各指标进行统计与方差分析,并以Duncan's新复极差法比较各时间段内指标间的差异显著性。
2 结果与分析
2.1 泽兰实蝇内生殖器官
泽兰实蝇的雌性内生殖器官主要由1对卵巢、输卵管、1对雌性附腺、1对受精囊等组成。卵巢左右成对,略呈椭圆形,基部通过侧输卵管连接到中输卵管上,羽化初期为淡黄色,后期变成黄色;附腺为一对无色透明囊状物,基部稍微膨大与输卵管相连;受精囊呈梨形或马蹄形,通过细长的受精囊管与雌性附腺和中输卵管相连(图1A)。
泽兰实蝇雄性生殖器官主要由1对精巢、5~7个大小不等呈囊状的雄性附腺及射精管组成。精巢为1对鲜黄色囊状物,通过透明、细长的输精管与射精管相连;雄性附腺透明,由5~7个气球形状的大小不等的腺体组成,末端也连接在射精管上;射精管是比输精管略粗的透明管(图1B)。
2.2 泽兰实蝇内生殖器官在成虫期内的生长发育
2.2.1 卵巢 由表1可以看出,雌性泽兰实蝇的卵巢长度在羽化初期显著增大(F=3.753;P=0.001),最大值出现在羽化后第4天;卵巢的最大宽度出现在羽化后第 1 天(F=4.017;P=0.001)。
2.2.2 雌性附腺 从表2可以看出,雌虫的附腺长度与最大宽度在羽化初期有增大的趋势,最大值均出现在羽化后第5天,但长度(F=1.214;P=0.2411)与宽度(F=1.488;P=0.0864)在各日龄间均没有显著差异。

图1 泽兰实蝇内生殖器官示意图Fig.1 Schematic diagram of internal reproductive organs in P.utilis
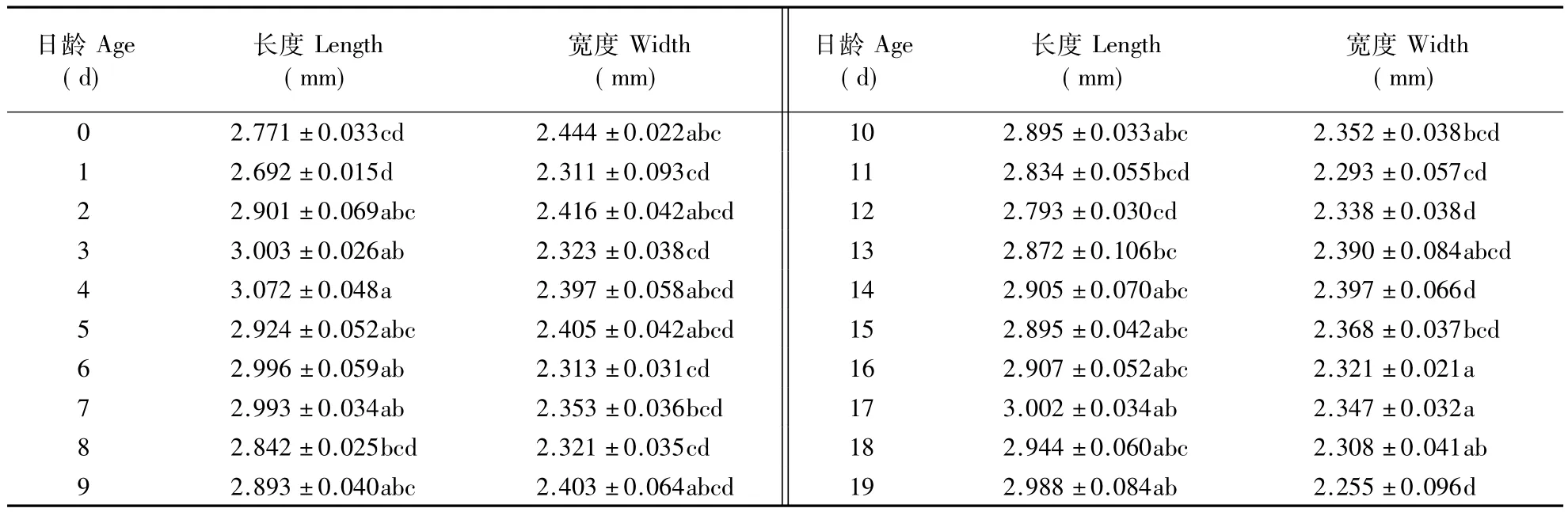
表1 不同日龄泽兰实蝇雌虫卵巢长度和宽度Table 1 Lengths and widths of female ovaries in P.utilis of different ages

表2 不同日龄泽兰实蝇雌虫附腺长度与宽度Table 2 Lengths and widths of female accessory gland in P.utilis of different ages
2.2.3 受精囊 在成虫发育过程中,受精囊长、宽见表 3,受精囊长度(F=1.084;P=0.365)和宽度(F=1.202;P=0.2518)的增长不明显,且各日龄间无显著差异。
2.2.4 精巢 不同日龄雄蝇的精巢长、宽如表4所示,雄蝇精巢长度在羽化初期显著增大(F=2.918;P=0.0001),最大值出现在第4天;精巢宽度在羽化后也显著增大,最大值出现于第2 天(F=11.553;P=0.0001)。2.2.5 雄性附腺 从表5可以看出,雄蝇附腺长度(F=1.054;P=0.3983) 与宽度(F=1.238;P=0.2258)在羽化初期均增长缓慢,长度在羽化后第4天达最大值,宽度的最大值出现在羽化后第5天。

表3 不同日龄泽兰实蝇雌虫受精囊长度与宽度Table 3 Lengths and widths of spermatheca in female P.utilis of different ages

表4 不同日龄泽兰实蝇雄虫精巢长度和宽度Table 4 Lengths and widths of male testes in P.utilis of different ages
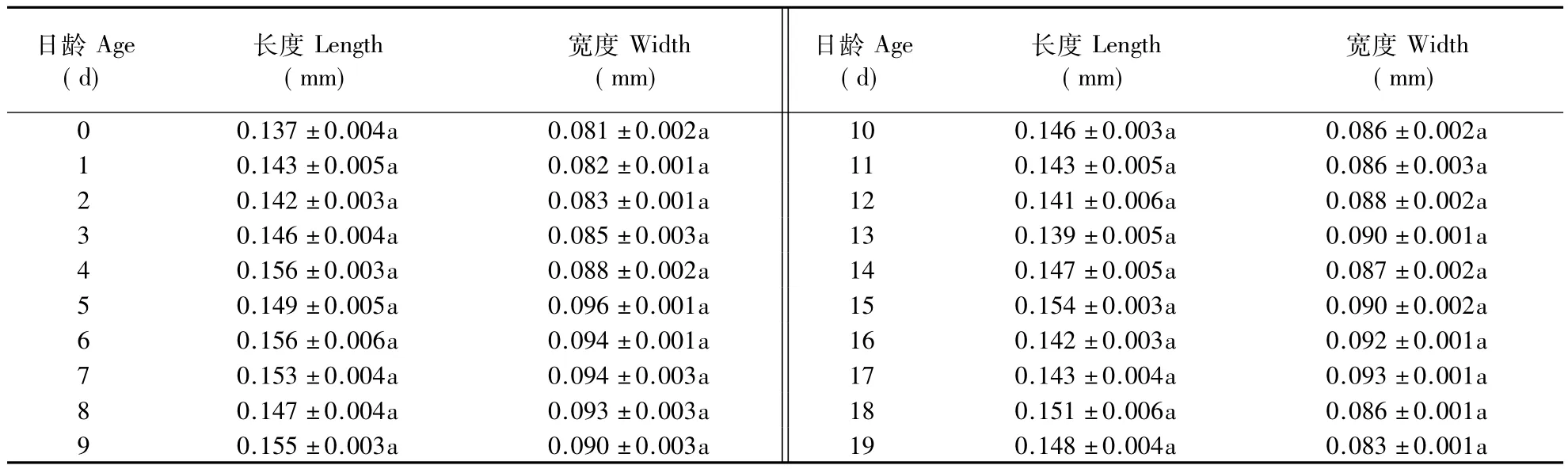
表5 不同日龄泽兰实蝇雄虫附腺长度与宽度Table 5 Lengths and widths of male accessory gland in P.utilis of different ages
3 讨论
泽兰实蝇的内生殖器官与其他蝇类昆虫较为相似,均可明显地分为卵巢、输卵管、受精囊、雌性附腺、生殖腔,精巢、输精管、雄性附腺、射精管等。然而,内生殖器官在蛹和成虫分类上极具参考价值(侯小燕等,2008)。有关蝇类雄性附腺的形态,目前已报道的几个科均不相同,如家蝇科没有雄性附腺(Matsuda,1976),麻蝇科的雄性附腺为1对较长的腺体(高熹等,2011),而本研究中观察到的泽兰实蝇的雄性附腺呈囊状,且个数较多,为5~7个。
本研究发现,泽兰实蝇卵巢长度的最大值出现在羽化后第4天,宽度最大值出现在第1天,且与其他日龄间有显著差异;雌性附腺、受精囊的大小在各日龄间均无显著差异;精巢长度在羽化后第4天出现峰值,而宽度在第2天出现峰值,且与其他日龄间有显著差异;雄性附腺的大小在各日龄间差异均不显著。由此可见,泽兰实蝇在羽化后主要生殖器官(卵巢、精巢)仍在增长,一般经过4 d的发育才有所停滞。然而,根据笔者观察及前人的报道,雌性泽兰实蝇在羽化后1 d左右可与雄性交配(陈升碧和关德盛,1991),这可能与羽化后第1天卵巢的宽度达最大值相关。有关交配日龄与产卵量的关系有待于进一步研究。
陈升碧,关德盛.1991.泽兰实蝇生物学特性观察及生物防治.西南农业学报,7(4):98-102.
陈旭东,何大愚.1990.泽兰实蝇对紫茎泽兰的控制作用及其评价研究.杂草学报,4(3):1-6.
高熹,曹凤勤,杨继雄,李强,吴国星.2011.棕尾别麻蝇雄性内生殖器官的结构及其发育.动物学研究,32(S):192-196.
何大愚,梁家社.1988.紫茎泽兰防治进展.生态学进展,5(3):163-168.
侯小燕,花保祯.2008.蝎蛉科(长翅目)昆虫雌性生殖系统构造及其在分类学上的意义.动物分类学报,33(3):427-434.
刘文耀,刘伦辉,和爰军.1991.泽兰实蝇对紫茎泽兰生长发育及生物量分配影响的研究.生态学报,11(3):21-24.
马沙,高熹,朱家颖,吴国星,汪金蓉,叶敏,肖春,李正跃.2012.温度及补充营养物对泽兰实蝇寿命的影响.生物安全学报,21(3):236-239.
唐启义,冯明光.2002.实用统计分析及其DPS数据统计分析系统.北京:科学出版社.
王晓芳.2010.中国黄粉蝶亚科昆虫雌性生殖系统研究(鳞翅目:粉蝶科).杨凌:西北农林科技大学.
魏艺,张智英,何大愚.1989.泽兰实蝇人工繁殖技术.生物防治通报,5(1):41-42.
詹光杰.2009.拟黑多刺蚁雌性生殖系统的比较观察.安徽农业科学,37(4):1578-1579,1620.
张智英,魏艺,何大愚.1988.泽兰实蝇生物学特性的初步研究.杂草学报,4(1):10-13.
郑征.1989.泽兰实蝇对紫茎泽兰生长发育及光合作用影响的研究.杂草学报,3(2):21-24.
周川江,周俗.2003.紫茎泽兰防治与利用概况.四川草原,(6):7-10.
Bennett P H and van Staden J.1986.Gall formation in crofton weed,Eupatorium adenophorumSpreng.(syn.Ageratina adenophora),by theEupatoriumgall flyProcecidochares utilisStone(Diptera:Trypetidae).Australia Journal of Botany,34:473-480.
Bess H A and Haramoto F H.1958.Biological control of pamadani,Eupatorium adenophorum,in Hawaii by a taphritid gall fly,Procecidochares utilispopulation studies of the weed,the fly and the parasites of the fly.Ecology,40:244 -249.
Davey K G.1985.The Male Reproductive Tract in Comprehensive Insect Physiology,Biochemistry and Pharmacology.New York:Plenum Press.
Dodd A P.1961.Biologocal control ofEupatorium adenophorumin Queensland.Australian Journal of Science,23:356 -365.
Matsuda R.1976.Morphology and Evolution of the Insect Abdomen.Oxford:Rergamon Press.
Rahman O and Agarwal M L.1991.Biological control of crofton weed(Eupatorinm adenophorumSpreng)by a fruit flyPocecidochares utilisStone in eastern Himalayas.Journal of Weed Science,22(1-2):98-101.
