水螅营养积累对基盘组织更新进程的影响
2013-07-05钱小成李晓飞刘正亮潘红春
钱小成 李晓飞 刘正亮 潘红春
(安徽师范大学生命科学学院,重要生物资源保护与利用研究安徽省重点实验室,生物环境与生态安全安徽省高校省级重点实验室,芜湖 241000)
淡水水螅是常见的两胚层低等动物,由于其具备在实验室易培养以及无性繁殖速度非常快等优点,因此水螅已成为在进化生物学、细胞生物学、发育生物学和动物再生等研究领域具有较高科研价值的模式生物[1,2]。水螅具有简单的单个体轴的极性体制,一个极为由口、垂唇和口周围的数根触手组成的口区,另一极为由柄和位于身体末端的基盘构成的足区,口区和足区之间为胃区,其中胃区与柄的分界处为出芽区。在野外自然水体中,水螅依靠基盘吸附于水中物体如水生植物的表面,其身体另外一端即水螅头部口周围的触手得以自然延长下垂“守株待兔”式地捕食水中快速运动的小型无脊椎动物,因此位于身体末端的基盘作为固着器官在水螅生命活动中发挥着重要的作用[3,4]。基盘组织的形态学、细胞组成及相关生理生化指标已较为明确[5-9],但基盘的固着机理及保持基盘组织的形态学完整性(基盘在附着和脱离水中支持物表面的过程中形态结构易受到破坏)的细胞学机制等方面仍未定论。
本研究室由于实验需要经常在高频喂食的条件下规模化培养水螅,在大批量水螅个体中偶然发现一些水螅个体出现更新基盘组织的现象,这个现象暗示水螅的营养积累可能是水螅基盘组织更新的诱因。基于此,本文为探讨水螅营养积累与基盘组织更新进程间的相关性,系统地观察了不同喂食频率条件下水螅更新基盘组织的进程,为进一步研究水螅基盘保持形态学完整性的细胞学机制提供基础资料。
材料和方法
1.实验材料
实验用水螅为本实验室培育的大乳头水螅(Hydramagnipapillata)单克隆无性繁殖系,原种采自安徽省芜湖市汀堂公园水塘(31。21’39.2”N,118。22’51.46”E)。实验前水螅每3d喂食一次卤虫幼体(Artemia sp.),喂食1h后更换培养液,然后维持在温度22±0.5℃的光照培养箱中(光照强度=2000LX,LD:HD=12h∶12h)。
2.实验方法
2.1 喂食频率与基盘更新进程相关性实验
挑取体长1.5cm左右且无芽体的水螅个体用于本实验。共分为5个实验组,每组的喂食频率分别为1天喂1次、1天喂2次、1天喂3次、1天喂4次和1天喂5次。所有实验在温度22±0.5℃、光照度2000LX以及光周期亮暗比12h∶12h的条件下进行,每个实验组20只水螅,每只水螅单独放在0.5L玻璃烧杯中培养,此后每隔24h通过Olympus数码显微镜观察和记录。每次喂食后均清污换水,所用水为除氯曝气并充氧的自来水。实验共进行了40d。
2.2 基盘分子标志物基盘过氧化物酶表达的检测
通过ABTS细胞化学染色法检测水螅基盘分子标志物过氧化物酶的表达,观察水螅基盘组织是否形成。ABTS细胞化学染色法的基本原理是反应液中的ABTS和H2O2均能一定程度地穿透细胞膜进入细胞质,在细胞质中表达的过氧化物酶可使H2O2分解为水和活性氧,而活性氧进一步氧化ABTS使之由还原态时的淡蓝色转变为氧化态时的紫红色,就这样表达过氧化物酶的细胞被染成紫红色[2-3]。将水螅置于白色陶瓷比色板上(每孔1只水螅),每孔分别加入350μl新鲜配制的ABTS染色混合液(655mmol/L柠檬酸、345mmol/L柠檬酸盐和1%ABTS溶液各700μl,70μl 0.3%H2O2,双蒸水定容至7ml)[4],反应20min后用双蒸水洗涤2次,加入300μl 2%NaF溶液,然后通过Olympus数码显微镜观察和拍照。
结 果
1.水螅更新基盘组织进程中的形态学变化
不同喂食频率下水螅更新基盘组织的时间进程虽有差异,但其形态学进程却类似(图1)。维持一定的喂食频率一段时间后,水螅在出芽区逐渐出现数个芽体(图1B-C),成熟芽体不断脱落,新生芽体不断出现,若干天后在出芽区和基盘之间靠近芽体的位置出现缢痕(图1D-F),最后水螅体在缢痕处断裂为水螅体主体和老基盘组织两部分(图1G)。
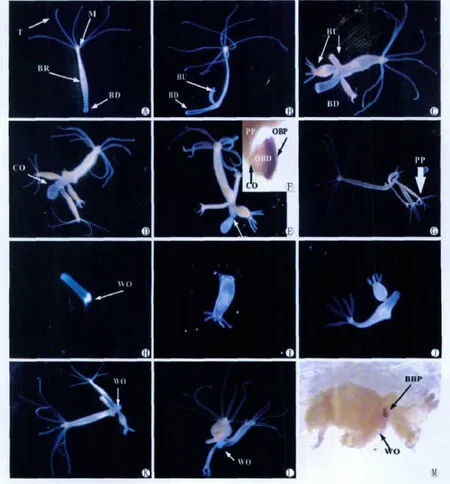
图1 1d喂3次实验组水螅更新基盘组织进程中的形态学变化.1d(A,10×),3d(B,10×),5d(C,10×),14d(D,10×),21d(E,10×),缢痕断裂前基盘过氧化物酶的表达(F,30×),23d时缢痕断裂(G,6×),缢痕断裂12h后的老基盘组织(H,15×),缢痕断裂48h后的老基盘组织(I,10×),缢痕断裂72h后的老基盘组织(J,10×),缢痕断裂3d后水螅体主体(K,10×),缢痕断裂15d后水螅体主体(L,10×),缢痕断裂15d后水螅体主体伤口处过氧化物酶表达的检测(M,20×),BBP(芽体基盘组织过氧化物酶的表达),BD(基盘),BR(出芽区),BU(芽体),CO(缢痕),M(口),OBD(老基盘组织),OBP(老基盘组织过氧化物酶的表达),PP(水螅体主体),T(触手),WO(缢痕断裂后的伤口)Fig.1 The morphological process of hydra renewing basal disc under the feeding frequency of thrice daily.1d(A,10×),3d(B,10×),5d(C,10×),14d(D,10×),21d(E,10×),the expression of peroxidase in the basal disc before the hydra broke into two parts at the constriction(F,30×),the hydra broke into two parts at the constriction after 23days under the feeding frequency of thrice daily(G,6×),old basal disc tissue at 12hafter the hydra broke into two parts at the constriction(H,15×),old basal disc tissue at 48hafter the hydra broke into two parts at the constriction(I,10×),old basal disc tissue at 72hafter the hydra broke into two parts at the constriction(J,10×),the parent polyp at 3dafter the hydra broke into two parts at the constriction(K,10×),the parent polyp at 15dafter the hydra broke into two parts at the constriction(L,10×),the detection of peroxidase expression in the wound of parent polyp at 15dafter the hydra broke into two parts at the constriction(M,20×),BBP(the expression of peroxidase in the basal disc of bud),BD(basal disc),BR(budding region),BU(bud),CO(constriction),M (mouth),OBD(old basal disc tissue),OBP(the expression of peroxidase in the old basal disc tissue),PP(parent polyp),T(tentacle),WO(wound)
2.喂食频率对基盘更新进程的影响
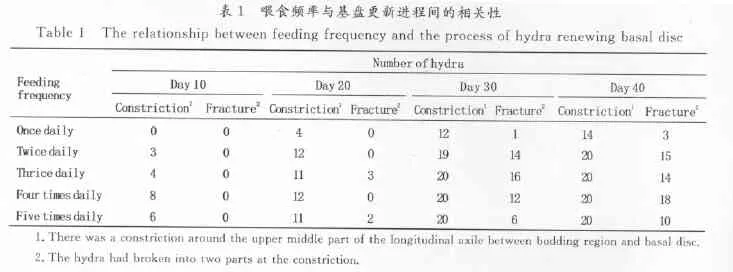
喂食频率对水螅基盘更新进程的影响较为明显(表1),实验进行到40d时,1天喂5次、1天喂4次、1天喂3次和1天喂2次实验组中的每个水螅个体在芽体下方均出现了缢痕,但1天喂1次实验组仅有70%的个体在芽体下方出现缢痕。另外,实验进行到40天时各实验组缢痕最终断裂的水螅个体数差异明显,1天喂4次、1天喂3次和1天喂2次实验组中70%以上的个体缢痕断裂,1天喂5次实验组50%的个体缢痕断裂,而1天喂1次实验组总共20只个体中只有3只缢痕断裂。
?
3.脱落的老基盘组织的两种命运
实验进行到40d时从所有实验组得到的共约60只脱落的老基盘组织在缢痕断裂后独立发育时呈现两种不同的命运,其中约1/3的老基盘组织的伤口再生出头部、直至发育成正常的水螅个体(图1H-J),其头部再生时间进程与水螅头部人工切除手术后头部再生时间进程基本相似[2];而另外约2/3的老基盘组织在缢痕断裂后独立发育时逐渐收缩和萎缩,其伤口处不能发育成头部,直至1w后解体(图2)。
4.老基盘组织脱落后水螅体主体新基盘组织的再生
缢痕断裂后对水螅体主体保持既定的喂食频率,其断裂伤口能愈合但不能再生出新的基盘组织(图1K-L),这个结论有两个重要依据,其一是愈合的伤口处(该部位本应再生出新生基盘组织)一直无固着能力;其二是愈合的伤口处未能检测出基盘特异性过氧化物酶(基盘分子标志物)的表达(图1M)。喂食频率降低到3天喂1次后,伤口上方的芽体逐渐脱落(图3C-F),直至其伤口上方的芽体全部脱落后愈合的伤口处再生出新生基盘组织(图3G-H)。
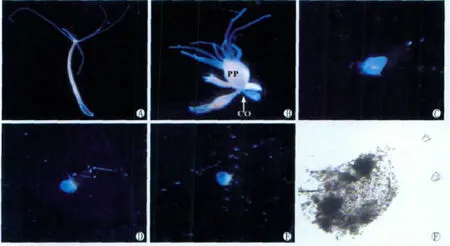
图2 1天喂3次实验组1例水螅个体更新基盘组织进程中的形态学变化.1d(A,10×),5d(B,15×),缢痕断裂12h后的老基盘组织(C,20×),缢痕断裂4d后的老基盘组织(D,20×),缢痕断裂6d后的老基盘组织(E,20×),缢痕断裂8d后的老基盘组织已分解(F,200×),CO(缢痕),PP(水螅体主体)Fig.2 The morphological process of hydra renewing basal disc under the feeding frequency of thrice daily.1d(A,10×),5d(B,15×),old basal disc tissue at 12hafter the hydra broke into two parts at the constriction(C,20×),old basal disc tissue at 4dafter the hydra broke into two parts at the constriction(D,20×),old basal disc tissue at 6dafter the hydra broke into two parts at the constriction(E,20×),old basal disc tissue at 8dafter the hydra broke into two parts at the constriction(F,200×),CO(constriction),PP(parent polyp)
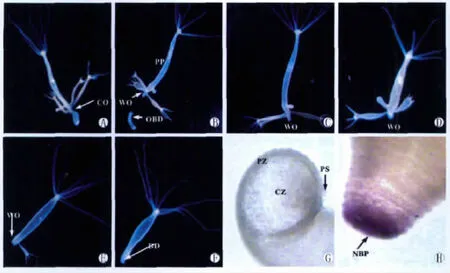
图3 缢痕断裂后水螅体主体伤口处新基盘组织的再生.1天喂3次进行到16d(A,10×),1天喂3次进行到24d(B,10×),缢痕断裂后降低喂食频率(降低到3天喂1次)第2d水螅体主体(C,10×),缢痕断裂后降低喂食频率第3d水螅体主体(D,10×),缢痕断裂后降低喂食频率第4d水螅体主体(E,10×),缢痕断裂后降低喂食频率第6d水螅体主体(F,10×),缢痕断裂后降低喂食频率第6d水螅体主体再生的新基盘组织吸附在玻片表面的形态(G,300×),缢痕断裂后降低喂食频率第6d水螅体主体再生的新基盘组织过氧化物酶的表达(H,250×),BD(基盘),CO(缢痕),CZ(不与玻片表面接触的基盘中央区域),NBP(新基盘组织过氧化物酶的表达),OBD(老基盘组织),PP(水螅体主体),PS(基盘组织周边区域伸出的伪足),PZ(基盘组织固着于玻片表面的周边区域),WO(缢痕断裂后的伤口)Fig.3 New basal disc regeneration of parent polyp after the hydra broke into two parts at the constriction.16dunder the feeding frequency of thrice daily(A,10×),24dunder the feeding frequency of thrice daily(B,10×),2dunder the lower feeding frequency of once every three days(C,10×),3dunder the lower feeding frequency of once every three days(D,10×),4dunder the lower feeding frequency of once every three days(E,10×),6dunder the lower feeding frequency of once every three days(F,10×),new basal disc attaching to glass slide(G,300×),the expression of peroxidase in the new basal disc of parent polyp(H,250×),BD(basal disc),CO(constriction),CZ(central zone of new basal disc),NBP(the expression of peroxidase in the new basal disc of parent polyp),OBD(old basal disc),PP(parent polyp),PS(pseudopod),PZ(perimeter zone of new basal disc),WO(wound)
讨 论
研究发现的淡水水螅更新基盘组织的特殊现象以前在刺胞动物门水螅纲动物中未见报道,其生物学意义值得深入探讨[10]。从水螅体本体脱落的老基盘组织的两种不同的命运(大部分的老基盘组织不能发育成正常水螅体、最终解体;而小部分的老基盘组织能发育成正常水螅体)折射出水螅更新基盘组织的生物学意义可能是多元的。在自然水体中水螅凭借其基盘附着在水中的植物或其他物体的表面,同时基盘表面上皮细胞还可伸出伪足而使身体沿着支持物慢慢移动,有时基盘也可脱离原来的支持物、进而转移到其他支持物上[4]。水螅基盘在附着、脱离支持物的过程中其细胞结构容易受到损伤,这可能是水螅更新基盘组织的内在动因。
然而,实验结果表明,也有一部分从水螅体本体脱落的老基盘组织正常发育成新生水螅,似乎水螅更新基盘的现象是一种无性生殖形式,特别是水螅断裂前在出芽区和基盘之间靠近芽体位置出现的缢痕(图1D-F)提示这种更新基盘的过程是一种无性横裂生殖。较低喂食频率(1天喂1次)实验组缢痕最终断裂的水螅个体数量很少、而其他实验组(1天喂2-5次)缢痕最终断裂的水螅个体数量较多(表1),喂食频率其实代表水螅营养积累水平[11],因此本文实验结果实际上反映的是较高营养积累水平能促进水螅更新基盘组织。较高营养水平首先促进水螅出芽生殖(图1C,图3A),在出芽区下方缢痕产生之前出芽区一直保持数个芽体(存在的芽体是动态的,成熟芽体不断脱落,新生芽体不断出现),这种高频率的无性出芽生殖持续一段时间后在出芽区下方开始出现缢痕。水螅个体大小是一定的,高营养积累水平刺激细胞分裂,最终以出芽生殖的形式保持水螅体内营养平衡,因此高频率的无性出芽生殖维持相当长时间后在出芽区和基盘之间靠近芽体的位置横断为前、后两个部分(水螅体本体及老基盘组织)可能是水螅在较高喂食频率下保持体内营养平衡的另外一种补充方式。总之,本文首次观察到的淡水水螅在出芽区和基盘之间的横断现象可以归纳为除出芽生殖外的第二种无性生殖方式,并且较高营养积累水平能刺激这种无性横裂生殖的发生。
另外,研究还发现缢痕断裂后对水螅体主体保持既定的喂食频率,其断裂伤口能愈合但不能再生出新的基盘组织;对其降低喂食频率直至其伤口上方的芽体全部脱落后断裂伤口处才开始启动产生新的基盘组织,这说明伤口附近的芽体抑制了该处新生基盘的再生。无独有偶,金灿等通过人工手术在水螅芽体下方紧贴芽体的位置把水螅切割为前后两个部分[12],手术后身体前段伤口处不能马上启动基盘再生机制,即使芽体脱离母体后仍然有相当数量的身体前段伤口不能再生出新生基盘。芽体抑制伤口处基盘组织再生的机制值得进一步探讨。
[1]Kasuminga O.Developmental neurobiology of hydra,a model animal of cnidarians.Canadian Journal Zoology,2002,80:1678-1689
[2]金灿,潘红春.水螅芽体发育与头部再生进程间的相互作用.中国组织化学与细胞化学杂志,2011,20(1):83-91
[3]Ghaskadbi SS,Shetye L,Chiplonkar S,et al.Ultraviolet irradiation initiates ectopic foot formation in regenerating hydra and promotes budding.Biosci,2005,30(2):177-182
[4]徐金金,潘红春,金灿等.大乳头水螅基盘再生进程中过氧化物酶的表达.中国组织化学与细胞化学杂志,2010,19(3):269-273
[5]Siebert S,Thomsen S,Reimer MM,et al.Control of foot differentiation in Hydra:Phylogenetic footprinting indicates interaction of head,bud and foot patterning systems indicates interaction of head,bud and foot patterning systems.Mechanisms of Development,2005,122:998-1007
[6]Leontovich AA,Zhang J,Shimokawa K,et al.A novel hydra matrix metalloproteinase(HMMP)functions in extracellular matrix degradation,morphogenesis and the maintenance of differentiated cells in the foot process.Development,2000,127,907-920
[7]Sabine H,Schaller HC.A new biochemical marker for foot-specific cell differentiation in hydra.Roux's Dev Biol,1985,194:453-46
[8]Hoffmeister-Ullerich SAH,Herrmann D,Kielholz J,et al.Isolation of a putative peroxidase,a target for factors controlling foot-formation in the coelenterate hydra.Eur J Biochem,2002,269,4597–4606
[9]Thomsen S,Bosch TCG.Foot differentiation and genomic plasticity in Hydra:lessons from the PPOD gene family.Dev Genes Evol,2006,216:57-68
[10]Hunter T.The energetics of asexual reproduction:Pedallaceration in the symbiotic sea anemone Aiptasia pulchella(Carlgren,1943).Journal of Experimental Marine Biology and Ecology,1984,83(2):127-147
[11]O.Chomskya,Y.Kamenira,M.Hyamsa,et al.Effects of feeding regime on growth rate in the Mediterranean Sea anemone Actinia equina (Linnaeus).Journal of Experimental Marine Biology and Ecology,2004,299:217-229
[12]金灿.大乳头水螅基盘特异性过氧化物酶细胞化学染色方法的建立及应用、编码基因克隆及原核表达.安徽师范大学硕士学位论文,2011,35-40
