GDF-9 和 BMP-15 在不同品系小鼠中的多态性和生殖能力关联分析
2013-05-22殷昆仑张长勇王天奇付淑霞何嘉玲孙德明
殷昆仑,张长勇,王天奇,暴 国,付淑霞,何嘉玲,孙德明
(1.北京协和医学院研究生院,北京 100730;2.国家人口计生委科学技术研究所,北京 100081)
生长分化因子9(growth differentiation factor 9,GDF-9)和骨形态发生蛋白15(bone morphogenetic protein 15,BMP-15)是由卵母细胞分泌的两种生长因子,对哺乳动物的正常生殖能力是不可或缺的[1]。GDF-9和BMP-15均属于转录生长因子β(transforming growth factor beta,TGFβ)超家族蛋白中的成员,其中GDF-9是第一个被证实与卵泡发育相关的基因。在小鼠,缺失基因GDF-9时卵泡发育被阻止在早期阶段而导致不孕[2],此后不久,在绵羊中多种BMP-15基因突变被认为是潜在的导致卵泡发育改变和不孕的因素[3,4]。目前许多研究已经证明GDF-9是控制绵羊高产仔的主效基因之一,Hanrahan等[4]将 Belclare绵羊和 Cambridge绵羊 GDF-9 基因编码区1184bp处的碱基突变(C→T)命名为G8突变,将BMP-15718处碱基突变718处碱基突变(C→T)命名为B2突变,同时将Belclare绵羊的BMP-15基因1100处的碱基突变(G→T)命名为 B4突变。Hanrahan等[4]发现 X 染色体上 BMP-15基因的突变和5号染色体上GDF-9基因的突变,与高繁殖力绵羊品种Belclare和Cambridge杂合携带母羊的高排卵数和纯合子不育相关联。GDF-9在小鼠定位于11号染色体的 29.0cM 处[5],全长 4618bp,包含两个外显子,其中外显子1的编码区是397bp,外显子2的编码区是926bp,共同编码一种由441个氨基酸组成的蛋白质,其成熟C-末端片段由135个氨基酸残基组成,与绵羊的同源性为66%[6]。而 BMP-15定位于 X染色体的2.81 cM,全长6618bp,同样也是两个外显子,其中外显子1的编码区是322bp,外显子2的编码区是857bp,共同编码一种由392个氨基酸组成的蛋白质。其特性与GDF9相似。本研究以高繁殖力小鼠(ICR)和低繁殖力小鼠(BALB/c)为实验材料,研究基因GDF-9和BMP-15在两种小鼠中的多态性,旨在找出与小鼠高繁殖力相关的SNP,为小鼠高繁殖力的标记辅助选择和育种提供理论依据。
1 材料
1.1 实验动物
ICR 小鼠,SPF 级(specific-pathogen free,SPF),雌雄各20只,雌鼠6周龄,雄鼠8周龄,由北京维通利华实验动物技术有限公司提供,许可证号【SCXK(京)-2010-0002】。BALB/c 小鼠,SPF 级,雌雄各20只,雌鼠6周龄,雄鼠8周龄,由北京华阜康生物科技股份有限公司提供,许可证号【SCXK(京)-2009-0007】。并按实验动物使用的3R原则给予人道的关怀。
1.2 主要试剂
全血基因组提取试剂盒、Taq酶、dNTPs(10mM)、DNA Marker2k均购自北京全式金生物技术有限公司;内切酶KpnI、XhoI购自宝生物工程(大连)有限公司;琼脂糖凝胶回收试剂盒购自天根生化科技(北京)有限公司;Dual-Luciferase® Reporter Assay System剂盒(Promega公司,美国);Lipofectamine® 2000 Transfection Reagent(Invitrogen公司,美国);测序由北京六合华大基因科技股份有限公司完成。
1.3 方法
1.3.1 小鼠繁殖
购回实验动物动物为SPF级别,适应饲养两周及实验均在 SPF级动物室进行【SYXK(京)2009-0033】,每一品系雌雄各20只随机配对,以产仔5胎来计算平均生殖能力,具体统计指标包括产仔数、离乳数和胎间隔时间。
1.3.2 小鼠全血基因组DNA提取
在同一时期对不同组别的每只小鼠进行尾静脉采血,用EDTA抗凝,并使用全式金基因组试剂盒提取基因组DNA,用去离子水溶解,测浓度和OD值后,琼脂糖凝胶电泳,保存于-20℃。
1.3.3 引物设计和 PCR扩增
根据 GeneBank发表的 GDF-9(登录号 NC_000077.6)和 BMP-15(登录号 NC_000086.7)基因的两个外显子的序列设计引物,引物序列及条件退火温度见表1。PCR反应总体积 25μL,其中 DNA为1μL(约 30ng),上下游引物终浓度为 200ng/μL,dNTPs为 0.5μL(终浓度为 200nM/μL),20mM Tris-HCl,pH8.3,20mM KCl,10mM(NH4)2SO4,2mM MgSO4,Taq DNA聚合酶 2.5U。反应条件:97℃预变性 5min,95℃ 变性 30s,65℃ 或者 63℃ 退火 30s,72℃延伸50s,35个循环;最后再 72℃延伸 5min,4℃保存。产物用1%的琼脂糖凝胶电泳检测。
1.3.4 PCR产物直接测序和分析
PCR产物测序使用的是ABI3730平台正向或反向测序,测序结果使用 DNAstar和 Chromas软件分析。
1.3.5 不同基因型的 GDF-9二级结构分析
对存在基因序列位点改变,分别使用 UCL和RNAfold软件预测位点变化分别对编码区和非编码区的二级结构的影响。

表1 扩增基因BMP-15和GDF-9外显子的引物Tab.1 Oligonucleotide primers for the amplification of the exons of the BMP-15 and GDF-9 exons
1.3.6 GDF-9双荧光素酶报告基因表达载体的构建
分别以ICR和BALB/c小鼠的全血基因组DNA为模板,扩增目的DNA片段,引物序列如下:
KpnI-F:5'GGGGTACCATGTACAGCTAGGCATG CTTGA 3';
XhoI-R:5'CCCTCGAGTTTCACTGGCTCCACT CTGGGATT 3'。PCR产物用内切酶 KpnI、XhoI进行酶切,酶切产物用天根凝胶回收试剂盒回收,回收的目的片段用 T4连接酶连接到 pGL3-Basic载体上,25℃反应15 min,构建重组质粒;并转化大肠杆菌DH5α菌株。酶切鉴定后测序验证。分别取100 μL测序验证正确的重组质粒加入到50 mL的LB培养基中,37℃、250 r/min转恒温晃动培养过夜,然后分别提取质粒DNA。
1.3.7 细胞的培养和转染
将293T细胞接种到48孔板进行培养,当细胞的密度达到90%时,使用1.3.6中提取的质粒,按照载体pGL3与pRL-TK载体6∶1的比例进行共转染,脂质体2000作为转染试剂使用脂质体2000进行共转染细胞。每组做3个复孔,实验平行重复3次。
1.3.8 双荧光素酶活性检测
转染48h后用双荧光素酶检测试剂盒进行检测:弃去孔内的培养基,用PBS洗一遍,然后每孔加入65μL的细胞裂解液,室温缓慢震荡15 min;加入30μLLARⅡ,检测萤火虫荧光素酶活性;再加入30 μL终止反应液,检测海肾荧光素酶活性。每种基因型一次做3个重复孔,结果表示为相对的荧光素酶活性(萤火虫荧光素酶活性/海肾荧光素酶活性)。实验平行重复3次。
2 结果
2.1 小鼠繁殖5胎的统计学分析
两种品系小鼠分别记录5胎繁殖记录,期间死亡的小鼠按数据缺失计;数据以均数±标准差(±s)表示;组间 t检验,p≤0.05为差异有显著性;使用SPSS13.0统计软件进行统计学分析,结果见表2。
表2 表2小鼠繁殖的记录¯±s)Tab.2 The mice breeding of record

表2 表2小鼠繁殖的记录¯±s)Tab.2 The mice breeding of record
注:对检测指标进行同列比较,A表示两组之间差异极显著(P<0.01),a表示两组之间差异显著(P<0.05).Note:Detect indicators of the same column,A represents the two groups was highly significant(P < 0.01),the a said between the two groups was significantly different(P <0.05)
胎间隔时间(天)a Tire interval ICR 20 5 100 14.25 ± 3.866 12.85 ±4.235 26.68 ± 8.186 BALB/c 20 5 100 5.01 ±2.556 3.80 ±3.012 31.06 ±1组别Group亲代数(对)Numbers of P胎数(胎)Tires样本量(例)Samples产仔数(只)A Litter size离乳数(只)A Weanling number 3.010
2.2 小鼠全血基因组DNA电泳图
注:泳道1为Marker2000,泳道2、3为 ICR品系;泳道4、5为 BALB/c品系。图1 小鼠基因组DNA琼脂糖凝胶电泳检测结果Note:Lane 1:D2000Marker;Lanes 2、3:ICR strain;Lanes4、5:BALB/c strain.Fig.1 Electrophoretic pattern of Mouse genome DNA
2.3 PCR产物测序结果
GDF-9在同一品系之间测序结果没有变化,但在不同品系之间碱基存在差别,在外显子1的上游非编码区的第8个碱基处,ICR为 T,BALB/c为 C(图 2-A、a,彩插 1);外显子 2 的 490 位 ICR 为 C,BALB/c 为 T(图 2-B、b,彩插 1),这不引起氨基酸的变化;746 位 ICR 为 T,BALB/c为 C(图 2-C、c,彩插1),该点在ICR为甲硫氨酸,在BALB/C为苏氨酸。其中甲硫氨酸和苏氨酸均为极性中性氨基酸,甲硫氨酸是含硫必需氨基酸,为含硫 α-氨基酸之一,是哺乳动物的必需氨基酸和生酮氨基酸。其侧链易氧化成甲硫氨(亚)砜,在α螺旋结构中更常见。苏氨酸含有一个醇式羟基的脂肪族α氨基酸,L-苏氨酸是组成蛋白质的20种氨基酸中的一种,有两个不对称碳原子,可以有4种异构体,是哺乳动物的必需氨基酸和生酮氨基酸。BMP-15不论是同一品系还是不同品系、组内或者组间,序列均没有的变化。
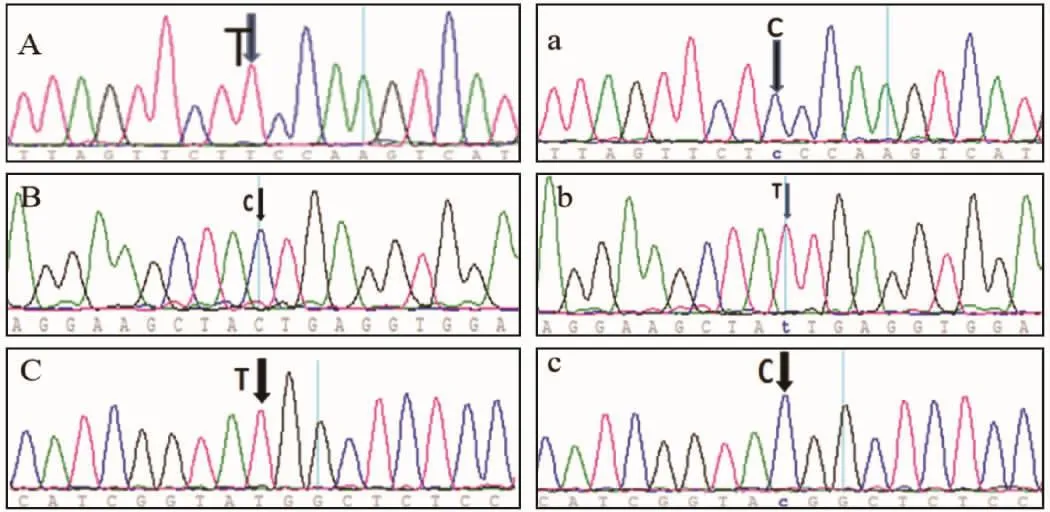
注: 图 A、B、C 代表 ICR 品系,图 a、b、c代表 BALB/c品系图2 GDF-9外显子测序结果Note: Figure A, B, C, representative of ICR strain, Figure a, b, and c represent the BALB / c strainFig.2 GDF-9 exon sequencing results
2.4 GDF9二级结构预测分析
为了验证序列变化对非编码区和编码区二级结构的影响,我们分别使用RNAfold和UCL在线预测不同基因型的 GDF-9的二级结构。非编区位点变化没有引起GDF-9二级结构的变化,但对于其结构能量熵有一定的影响,由此可知,该位点变化时对其结构的稳定性有一定的影响。编码区氨基酸的变化对其二级结构的影响较小,为只有ICR的α螺旋结构比BALB/c稍长(见图3、4)。
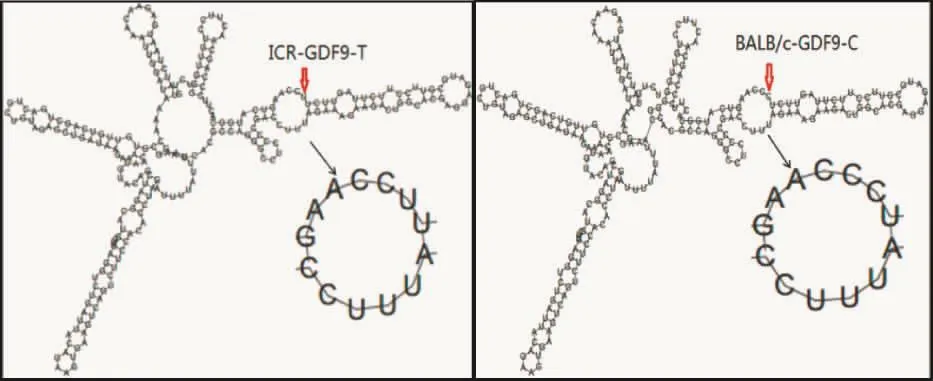
图3 GDF-9外显子1上游第8位碱基变化对非编码区二级结构的影响Fig.3 The eighth base change of the GDF-9 exon 1 impact on non-coding region secondary structure influence

注:左边为ICR品系,右边为BALB/c品系。图4 GDF-9外显子2序列变化对蛋白二级结构的影响Note:The left is ICR strain,and the right is BALB/c strain.Fig.4 GDF-9 exon 2 sequence change on the influence of protein secondary structure
2.5 载体构建示意图
我们将有变化的5’UTR连接到 pGL3-Basic载体上,由于Basic载体上没有启动子,所以若该区域具有启动子的功能则下游萤火虫基因就会表达,构建的载体示意图见图5。
2.6 双荧光素酶相对活性检测结果:
由双荧光素酶活性可以看出,BALB/c品系中GDF-9上调下游萤火虫荧光素酶基因的表达能力比ICR 低 38.4% 左右,P < 0.05(P=0.001)具有统计学差异。可以看出 GDF-9通过非编区影响 GDF-9基因的表达从而对于小鼠的繁殖能力是有一定的作用。

图5 载体结构示意图Fig.5 Carrier structure diagram
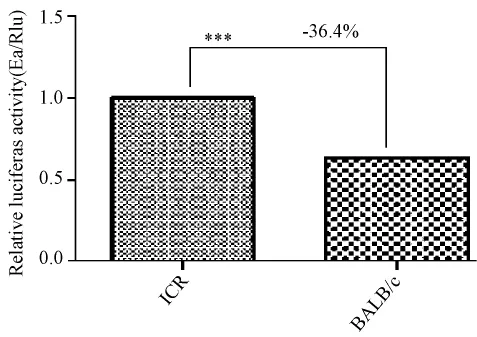
图6 双荧光素酶相对活性检测Fig.6 Dual luciferaserelative activitydetection
3 讨论
3.1 ICR和BALB/c小鼠繁殖能力的比较
本研究之所以选择一个近交系和一个远交系品种,是考虑到这两个品系之间在产仔数目上存在明显的差别,我们目的就是要研究除了自身品系之间的差别外,在基因水平是否存在能够区分不同繁殖能力的标记性的SNP位点。对不同品系的小鼠产仔能力进行了比较,与孟琼等[7]人研究相比,ICR产仔数目较以往有所提高,ICR产仔数目是BALB/c的2.8倍左右。
3.2 GDF-9基因的多态性
与以往的研究相比,本研究是第一次发现了ICR和BALB/c小鼠在 GDF-9中的5’UTR和编码蛋白成熟区存在多态性位点,5’UTR ICR-T对下游基因的表达起调节作用。在两种品系小鼠 GDF-9中发现了三个基因多态性位点,一个位于5’端非编码区 ICR到 BALB/c的碱基变化(T→C),;另外两个位于成熟的蛋白编码区,其中一个引起了氨基酸的变化,另一个是无义变化。但单独在两种品系中均没有检测到SNP位点。
另外,本研究测序所得的ICR品系小鼠的GDF-9基因第一个外显子和第二个外显子与Gene Bank所公布的C57小鼠的基因序列同样也存在3个碱基的不同,这种差异可能是由于小鼠品系不同造成的。
3.3 GDF-9的不同基因型的二级结构变化
本研究运用分子生物学知识预测了不同基因型的GDF-9的二级结构。对于研究 GDF-9在繁殖中的作用开辟了新途径,即除了编码区的作用之外,非编码区的不同也对基因的功能起一定的调控作用。
3.4 GDF-9和BMP-15基因与繁殖力的关系
GDF-9是通过旁分泌方式对卵泡的生长和分化起重要作用[8]。在小鼠中,除原始卵泡以外的各个时期卵泡的卵母细胞中,GDF-9 mRNA及蛋白持续表达至排卵后,于受精后1.5天消失。利用重组GDF-9蛋白在体外研究证实:GDF-9在卵泡生长和发育的各个阶段均有生物学活性并重要作用。
GDF-9在卵泡发育早期阶段的重要性在一些哺乳动物中已有得到证实,例如 GDF-9缺陷小鼠[2]、母羊的自发纯合突变[4]或免疫接种抵御 GDF-9[9]都表现出阻滞卵泡处于早期阶段。McIntosh等通过主动免疫小鼠-为 GDF-9和 BMP-15的前体蛋白对卵母细胞从卵巢分泌出后起重要的监管作用提供了生理学上的证据,包括控制排卵率和产仔数目[10]。在体实验表明 GDF-9也会造成原始卵泡数量的减少[11],体外暴露啮齿类动物[12]和人类[13]的卵巢组织促进了初级卵泡的进程。
我们的研究得到 GDF-9在不同繁殖力的小鼠中,高繁殖力的ICR品系中基因型不同的5’UTR比BALB/c可以更有效的促进 GDF-9的表达,通过双荧光素酶报告分析可以给我们以下的指示:产仔能力的大小可能与 GDF-9的表达量之间存在剂量依赖作用。这一点AlonKedem等的研究发现,从女性得到的初级卵母细胞对于重组的GDF-9和BMP-15存在剂量依赖的方式,将分离的卵母细胞暴露在这两种生长因子中可以增加发育中的卵母细胞的数量[14]。同时 Janet和 Kenneth 研究发现 GDF-9 和BMP-15的表达水平比例在每个物种都不相同,这点也支持了GDF-9和BMP-15的mRNA水平的不同是调控排卵率表型的关键因素[15]。
3.5 造成BALB/c繁殖能力降低的原因
目前,对于小鼠生殖力研究大多是在生理机制方面展开的,从现有文献中还没有看到与基因多态性相关的报道,本研究证明了在ICR和 BALB/c小鼠中GDF-9存在基因多态性位点,该位点与小鼠的繁殖力有密切的关系。影响小鼠繁殖力的因素包括遗传、环境、营养、管理和疾病等五大因素。然而,在小鼠被生物净化并在标准化的饲养环境条件下,环境、营养、管理和疾病等因素被严格地控制,而遗传特性则成为影响小鼠繁殖力最本质和内在的重要因素[16]。
其中,近交衰退是影响BALB/c繁殖能力低下的一个主要原因。从遗传学角度来看主要可以从以下两方面进行解释:1.有害的隐性基因的暴露。一般病态的突变基因绝大多数都是隐性的,所以处于杂合状态时不表现出病态或不利的性状。这些有害基因的作用可被显性的杂合子等位基因所掩盖,但经过一段近亲繁殖,纯合的基因(纯合子)比例渐渐增多,于是有害的隐性基因相遇成为纯合子而显出作用,出现不利性状,对个体的生长发育、生活和生育等产生明显的不利影响。2.多基因平衡的破坏。个体的发育受多个基因共同作用的影响,虽然其中每个基因的作用效应微小。对环境适应较好的野生或杂交动物,由于自然选择的作用有利于保存那些生物适应能力较强的基因组合具有平衡的多基因系统,近交繁殖往往会破坏这个平衡,造成个体发育的不稳定[17]。
目前在环境毒理和人类避孕方面的研究中需要高繁殖力的小鼠,若能找出繁殖力与基因之间的关系,就可以利用提前筛选性培育技术来提高小鼠的生产繁殖效率,提高实验动物的经济效益,也可以为筛选相关的模型动物种群提供帮助。本研究发现了GDF-9在繁殖能力不同的品系小鼠 ICR和BALB/c中的多态性,为筛选相关的模型动物种群提供参考资料。
[1]Otsuka F,McTavish KJ,Shimasaki S.Integral role of GDF-9 and BMP-15 in ovarian function[J].MolReprod Dev2011,78:9-21.
[2]Dong J, AlbertiniD F, NishimoriK, etal.Growth differentiation factor-9 is required during early ovarian folliculogenesis[J].Nature,1996,383,531 -535.
[3]Galloway SM,McNatty KP,Cambridge LM,et al.Mutations in an oocyte-derived growth factorgene(BMP - 15)cause increased ovulation rate and infertility in a dosage-sensitive manner[J].Nat.Genet,25,279 - 283.
[4]Hanrahan J.P.,Gregan S.M.,Mulsant,et al.Mutations in the genes for oocyte-derived growth factors GDF-9 and BMP-15 are associated with both increased ovulation rate and sterility in Cambridge and Belclare sheep(Ovisaries)[J].Biol.Reprod,70,900 -909.
[5]http://www.informatics.jax.org/.
[6]Bodensteiner K J,Clay C M,Moeller C L,et al.Molecular cloning of the ovineGrowth/Differentiation factor-9 gene and expression of growth/differentiation factor-9 in ovine and bovine ovaries[J].Biol Report,1999,60(2):381 - 386.
[7]Kaivo-Oja N, Bondestam J, Kämäräinen M, et al.Growth differentiation factor-9 induces Smad2 activation and inhibin B production in cultured human granulosa-luteal cells [J].J ClinEndocrinolMetab,2003,88(2):755 - 762.
[8]孟琼,杨锡平,胡一江,等.3个不同品种小鼠繁殖性能及生长发育的比较观察[J]。湖南中医学院学报,2000,20(03):9-10.
[9]Juengel JL,Hudson NL,Berg M,et al.Effects of active immunization against growth differentiation factor 9 and/or bone morphogenetic protein 15 on ovarian function in cattle[J].Reproduction,2009,138(1):107 -114.
[10]C Joy McIntosh,Steve Lawrence,Peter Smith,et al.Active immunization against the proregions of GDF-9 or BMP15 alters ovulation rate and litter size in mice[J].Reproduction,2012 Feb,143(2):195 -201.
[11]Vitt UA,Hayashi M,Klein C,et al.Growth differentiation factor-9 stimulates proliferation butsuppresses the folliclesti mulating hormone-induced differentiation of cultured granulosa cells from small antral and preovulatory ratfollicles[J].BiolReprod,2000a,62(2):370 -377.
[12]Hayashi M,McGee EA,Min G,et al.Recombinant growth differentiation factor-9 (GDF-9) enhances growth and differentiation of cultured early ovarian follicles[J].Endocrinology,1999,140(3):1236-1244.
[13]Nilsson EE,Skinner MK.Growth and Differentiation Factor-9 Stimulates Progression of Early Primary but Not Primordial Rat Ovarian Follicle Development[J].Biology of Reproduction,2002,67:1018 -1024.
[14]AlonKedem, Benjamin Fisch, RoniGaror, etal.Growth Differentiating Factor 9 (GDF-9)and Bone Morphogenetic Protein 15 both Activate Development of Human Primordial Follicles in Vitro,with Seemingly More Beneficial Effects of GDF-9[J].J Clin Endocrinol MeTab,2011,96(8):E1246 -1254.
[15]Janet L.Crawford,Kenneth P.McNatty.The ratio of growth differentiation factor 9:Bone morphogenetic protein 15 mRNA expression is tightly co-regulated and differs between species over a wide range of ovulation rates[J].Molecular and Cellular Endocrinology.348(2012)339– 343.
[16]孙德明,李根平等.《实验动物从业人员上岗培训教材》[M].北京:中国农业大学出版社,2011:3-4.
[17]Charles worth D,Willis JH.The genetics of inbreeding depression[J].Nature reviews Genetics.Nov,2009,10(11):783 -796.
