RNA干扰CLEC2B基因沉默对淋巴细胞可溶性白介素2受体mRNA表达的影响
2013-04-20张峻岭柳君如徐士福马秀亮
张峻岭,柳君如,徐士福,程 琳,马秀亮
白癜风为一种自身免疫相关性皮肤病已成共识,研究发现白癜风患者皮损内杀伤性T淋巴细胞浸润可以直接破坏黑素细胞,近年发现,免疫系统失调与基因的差异表达密切相关。国外学者Yu等[1]应用基因芯片技术发现白癜风皮损区CLEC2B mRNA表达高于非皮损区。笔者应用实时荧光定量反转录聚合酶链式反应(RT-PCR)方法,发现白癜风患者皮损区CLEC2B基因较正常人表达增加,并发现CLEC2B基因过表达的淋巴细胞培养上清液对黑素细胞的增殖和黑素合成有一定的抑制作用[2],同时发现CLEC2B基因过表达可上调与白癜风密切相关的细胞因子可溶性白介素2受体(sIL-2R)mRNA的表达。为进一步阐明CLEC2B基因在白癜风发病中的作用,在此基础上本研究利用RNA干扰技术,使 CLEC2B基因沉默,观察CLEC2B基因沉默后淋巴细胞sIL-2R mRNA的变化,旨在探索该基因功能相关细胞因子及其参与的信号传导途径,从而推测该基因参与白癜风发病的免疫学机制。
1 材料与方法
1.1 主要试剂与材料 DMRIE-C脂质体及OPTI MEM购于美国Invitrogen公司,CLEC2B引物序列由不列颠哥伦比亚大学Youwen zhou教授惠赠,CLEC2B siRNA由上海吉玛制药技术有限公司设计合成,RPMI 1640液态培养基购于美国Gibco公司,DMEM高糖液态培养基、TBD胎牛血清购于天津灏洋生物科技有限公司,TRIzol、TRANScript cDNA第一链合成试剂盒、SuperReal PreMix(SYBR Green I)均购于天根生化科技(北京)有限公司,MTT细胞增殖及细胞毒性检验试剂盒购于南京凯基生物科技有限公司,DMSO购于美国Sigma公司,人Jurkat淋巴瘤细胞株来自中国科学院血液病研究所。
1.2 方法
1.2.1 细胞培养 细胞培养条件均为37 ℃,5%二氧化碳(CO2)进行培养,Jurkat淋巴瘤细胞用RPMI 1640培养基,以密度为2×105/ml接种于25 cm2培养瓶,待细胞处于对数生长期、密度近1×106/ml(融合度近80%)时进行实验。
1.2.2 siRNA转染 DMRIE-C脂质体转染3条CLEC2B siRNA入Jurkat淋巴瘤细胞,采用反向转染法[3],转染体系为600 μl,siRNA终浓度约80 nM,siRNA∶DMRIE-C实际配比=3 μl∶2 μl。实验共设定CLEC2B沉默组(沉默1组、沉默2组、沉默3组)、非沉默对照组(转染scrambled siRNA,该siRNA无基因干扰效应,是与目的基因序列无同源性的阴性对照siRNA序列)及空白对照组(不做任何处理),转染具体步骤按试剂盒说明操作。
1.2.3 CLEC2B基因沉默效率检测 转染后48 h,收集细胞,进行RNA提取、cDNA合成及实时荧光定量RT-PCR方法检测CLEC2B基因沉默效率。具体步骤按照试剂盒说明操作。CLEC2B上游引物5′-CCCCTATGATTGGATTGGTT-3′,下游引物5′-GGCATGTTGAGTGGAACAGT-3,产物大小79 bp;β-actin上游引物 5′-CCTGGAGAAACCTGCCAAGT-3′,下游引物 5′-TGGGAGTTGCTGTTGAAGTC-3′,产物大小124 bp。应用RG-3000荧光定量PCR仪(澳大利亚Corbett Research Sydney 公司)进行检测以选取沉默效率最高的CLEC2B siRNA进行后续实验。
1.2.4 CLEC2B基因沉默后淋巴细胞增殖率检测 实验分组:空白对照组(未进行转染)、非沉默对照组(转染scrambled siRNA)、CLEC2B沉默组(转染最佳CLEC2B siRNA),每组设3个复孔。选择基因沉默效率最高的CLEC2B siRNA进行转染,转染后培养48 h,MTT法测细胞增殖率,具体步骤按说明书进行。酶标仪在570 nm波长处测各孔的光密度(A值)。
1.2.5 sIL-2R mRNA表达水平 选择转染后48 h CLEC2B基因沉默率最高的转染组,进行RNA提取、cDNA合成及实时荧光定量PCR方法检测sIL-2R mRNA表达情况,具体步骤按照试剂盒说明操作。实验分为3组:空白对照组(未进行转染)、非沉默对照组(转染scrambled siRNA)、CLEC2B沉默组(转染最佳CLEC2B siRNA),每组设3个复孔。sIL-2R引物经Pubmed检索基因序列后,由南京金瑞斯生物科技有限公司设计合成。sIL-2R上游引物5′-GAATTTATCATTTCGTGGTGGTCTCCG-3′,下游引物5′-TCTTCTACTCTTCCTCTGTCTCCG-3′,产物大小298 bp,三步法PCR反应条件:95 ℃预变性15 min,95 ℃变性5 s,64 ℃退火20 s,72 ℃延伸30 s,共40个循环。
1.2.6 结果分析 基因表达相对变化采用2-△△Ct法处理数据[4]。统计分析时用△Ct代表基因的相对定量,Ct值代表目标基因扩增产物达到设定阈值所经历的循环数。△Ct=目的基因Ct值-内参基因Ct值,△△Ct=待测样本△Ct-标准样本△Ct。
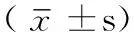
2 结果
2.1 CLEC2B基因沉默效率检测 转染CLEC2B siRNA后,各CLEC2B沉默组较非沉默对照组CLEC2B mRNA表达下降,差异均有统计学意义(P<0.05)。各CLEC2B沉默组siRNA序列对CLEC2B基因沉默效率不等,差异有统计学意义(P<0.01);其中沉默2组siRNA沉默效率最高,与沉默1组和沉默3组比较,差异均有统计学意义(P<0.05,见表1)。
2.2 CLEC2B基因沉默后对淋巴细胞增殖率及其sIL-2R mRNA表达的影响 CLEC2B基因沉默后,空白对照组、非沉默对照组、CLEC2B沉默组淋巴细胞增殖率比较,差异无统计学意义(P>0.05)。3组sIL-2R mRNA相对表达水平比较,差异有统计学意义(P<0.05);其中CLEC2B沉默组较非沉默对照组降低,差异有统计学意义(P<0.05,见表2)。

Table1 Comparison of CLEC2B mRNA expression quantity and slicensing effect among each group
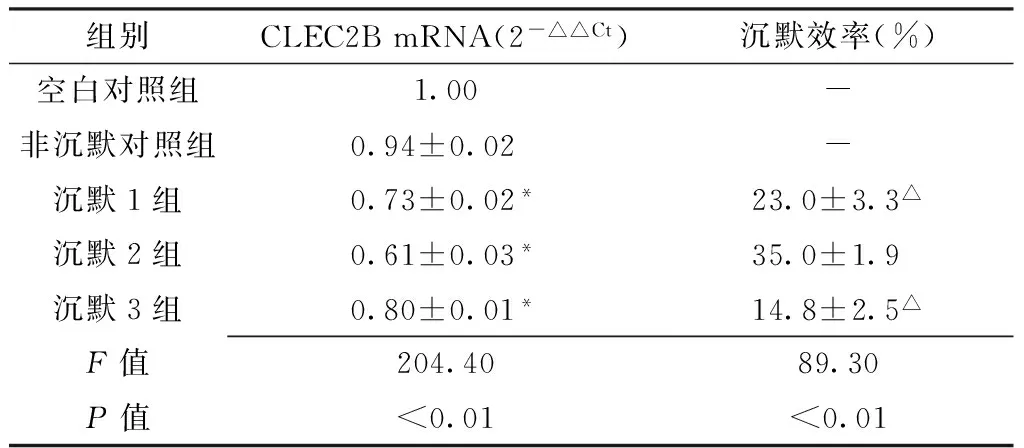
组别CLEC2BmRNA(2-△△Ct)沉默效率(%)空白对照组1.00-非沉默对照组0.94±0.02 -沉默1组0.73±0.02*23.0±3.3△沉默2组0.61±0.03*35.0±1.9 沉默3组0.80±0.01*14.8±2.5△F值204.4089.30P值<0.01<0.01
注:与非沉默对照组比较,*P<0.05;与沉默2组比较,△P<0.05
Table2 Comparison of lymphocytes proliferation rate and sIL-2R mRNA expression among each group

组别A值sIL-2RmRNA(2-△△Ct)空白对照组0.39±0.041.00非沉默对照组0.35±0.030.87±0.04 CLEC2B沉默组0.36±0.040.34±0.03*F值1.95435.80P值>0.05<0.01
注:与非沉默对照组比较,*P<0.05
3 讨论
CLEC2B基因编码位于12号染色体12p13-p12自然杀伤(NK)细胞基因复合体区,与NKG2D、 LOX-1和Dectin-1等构成复合体,属于C型凝集素超家族成员,有研究显示CLEC2B基因表达于各种血细胞[5],并可在活化的淋巴细胞中转录表达增加,为淋巴细胞活化后早期转录表达的基因[6]。本实验发现RNA干扰淋巴细胞中CLEC2B基因沉默后,细胞因子sIL-2R mRNA表达下调,反之可推测,白癜风患者体内CLEC2B的过表达可能引起sIL-2R水平升高,这对激活NK细胞杀伤活性、活化T淋巴细胞有重要意义。与此同时,国内外多项实验均证实IL-2、sIL-2R在白癜风患者血清中表达升高,且与白癜风严重程度相关[7],通过本实验结果进一步推测sIL-2R与CLEC2B参与的信号传导途径有一定相关性,参与白癜风的发病机制可能是:IL-2可诱导NK细胞增殖分化,亦可刺激NK细胞产生γ干扰素(INF-γ),反之INF-γ既能活化NK细胞,又能诱导NK细胞表达IL-2受体,从而增强细胞对IL-2的反应性。这样,就形成一个NK细胞-IL-2-INF-γ系统,既可以对NK细胞的数量和功能进行自我调节,又可以调节T、B淋巴细胞及其他免疫细胞的数量和功能[8-9]。已有研究表明,NK细胞与T淋巴细胞之间存在密切的免疫相关性,多种细胞因子参与彼此之间的调节,其共同终末效应为作用于黑素细胞,影响其存活及黑素合成功能。
本实验通过RNA干扰沉默CLEC2B基因,实验数据表明这直接影响了淋巴细胞sIL-2R的表达,结合本课题组既往已验证白癜风患者皮损内CLEC2B基因表达增多的现象,可推断CLEC2B基因参与白癜风发病机制可能通过影响sIL-2R等细胞因子转录表达,直接刺激T淋巴细胞活化分化和间接影响NK细胞的功能,最终引发黑素细胞减少白癜风的发生,证实了CLEC2B是参与白癜风发病的信号传导途径之一,而抑制该基因转录表达可阻止白癜风的病情进展,这为白癜风基因学及免疫学研究及白癜风的基因治疗提供了新的研究方向。目前,国内外对CLEC2B基因的研究多集中于抗肿瘤及抗感染等领域,而近期,CLEC2B参与的NK细胞相关免疫机制已受到国内外研究关注[10],关于该基因的功能及其他相关免疫学机制还有待继续深入研究。
1 Yu R,Broady R,Huang Y,et al.Transcriptone analysis reveals markers of aberrantly activated innate immunity in vitiligo lesional and non-lesional skin[J].PLoS One,2012,7(12):e51040.
2 张峻岭,徐士福,柳君如,等.CLEC2B基因过表达的Jurkat细胞培养上清液对黑素瘤细胞B16的影响[J].中国中西医结合皮肤性病学杂志,2012,11(6):348-350.
3 杨萍,严金川,刘培晶.siRNA反向转染法提高原代悬浮细胞转染效率的应用[J].江苏大学学报:医学版,2010,20(3):267-269.
4 Pfaffl MW.A new mathematical model for relative quantification in real-time RT-PCR[J].Nucleic Acids Research,2001,29(9):e45.
5 Hamann J,Montgomery KT,Lau S,et al.AICL:a new activation-induced antigen encoded by the human NK gene complex[J].Immunogenetics,1997,45(5):295-300.
6 Eichler W,Ruschpler P,Wobus M,et al.Differentially induced expression of C-type lectins in activated lymphocytes[J].J Cell Biochem Suppl,2001(Suppl 36):201-208.
7 Singh S,Singh U,Pandey SS.Serum concentration of IL-6,IL-2,TNF-α,and IFNγ in Vitiligo patients[J].Indian J Dermatol,2012,57(1):12-14.
8 Boyman O,Krieg C,Homann D,et al.Homeostatic maintenance of T cells and natural killer cells[J].Cell Mol Life Sci,2012,69(10):1597-1608.
9 Seki S,Nakashima H,Nakashima M,et al.Antitumor immunity produced by the liver Kupffer cells,NK cells,NKT cells,and CD8 CD122 T cells[J].Clin Dev Immunol,2011,29(11):868345.
10 Kuttruff S,Koch S,Kelp A,et al.NKp80 defines and stimulates a reactive subset of CD8 T cells[J].Blood,2009,113(2):358-369.
