我国沿海双唇形突海链藻的形态学研究
2013-04-19扬赵秋兰吕颂辉
李 扬赵秋兰,吕颂辉
(1. 华南师范大学生命科学学院, 广东省水产健康安全养殖重点实验室, 广东省高等学校生态与环境科学重点实验室,广州 510631; 2. 韩山师范学院陶瓷学院, 潮州 521041; 3. 暨南大学生命科学技术学院,
赤潮与水环境研究中心, 广州 510632)
我国沿海双唇形突海链藻的形态学研究
李 扬1赵秋兰1,2吕颂辉3
(1. 华南师范大学生命科学学院, 广东省水产健康安全养殖重点实验室, 广东省高等学校生态与环境科学重点实验室,广州 510631; 2. 韩山师范学院陶瓷学院, 潮州 521041; 3. 暨南大学生命科学技术学院,
赤潮与水环境研究中心, 广州 510632)
唇形突是海链藻中常见的一类突起, 其形态学特征和排列方式是重要的分类学依据。绝大多数海链藻只有一个唇形突, 两个唇形突的现象仅报道于极少数海链藻种类。我国关于双唇形突海链藻的报道较少。研究利用扫描电镜(SEM)技术, 对采自我国沿海海域的自然水样, 以及分离获得的单克隆培养藻株进行了形态学观察, 针对其中的海链藻属Thalassiosira Cleve种类开展了形态学的专题研究。报道了4个具有双唇形突的海链藻种类, 分别是有翼海链藻T. bipartita (Rattray) Hallegraeff、脆弱海链藻T. fragilis Fryxell、亨氏海链藻T. hendeyi Hasle & Fryxell和维斯吉思海链藻T. visurgis Hustedt, 其中有3个为我国新记录种类: 脆弱海链藻、亨氏海链藻和维斯吉思海链藻。对每个种类的形态学特征、生活习性和生态分布进行了描述, 提供了扫描电镜照片, 对相似种类的形态学特征进行了比较研究, 分析了唇形突的变化类型及其分类学价值。
海链藻; 双唇形突; 形态学; 中国沿海
海链藻是一类常见的浮游硅藻, 是硅藻中少数几个大属之一[1,2], 也是中心纲乃至硅藻门的模式生物,开展了诸如硅藻基因组、有性繁殖生活史、环境指示生物等多方面的研究, 是很好的研究材料[1,2]。另外,一些海链藻种类还是我国沿海重要的赤潮种[3—5]。
海链藻种类繁多, 种间的形态学差异细微。绝大部分种类的鉴定, 只有在电镜下才能准确进行[2,6]。关键的分类特征是支持突(Fultoportula)、唇形突(Rimoportula)、闭合突(Occluded process)的数量和分布位置[1,6,7]。唇形突是海链藻中普遍存在的突起类型, 绝大部分海链藻都只有一个唇形突, 具有两个唇形突的现象仅报道于极少数海链藻种类。我国关于双唇形突海链藻的报道较少。本文报道了4种双唇形突海链藻, 其中3种为我国新记录种类。对其形态学特征进行了详细说明, 并对相似种类的形态学特征进行了比较研究。
1 材料与方法
1.1 材料
用小体积柱状采水器在布设于长江口和广东沿海的多个站位采集海水2 L, 每个样品采集两份水样, 其中一份现场用鲁哥氏液(Lugol’s solution)固定。另一份样品则用于单克隆藻株的分离和纯化。待藻株纯化之后, 取较高细胞密度的藻液50 mL左右, 同样用鲁哥氏液固定。单克隆藻株保存于光照培养箱内, 培养条件是20℃、12∶12光循环、光照强度约为50—80 μmol photons/(m2·s)。
1.2 方法
固定后的水样经过多次的静置、沉淀, 逐步浓缩到5—10 mL。在浓缩后的样品中加入等体积的浓硫酸(浓盐酸), 并在水浴中煮沸15—20min左右, 具体时间可根据有机质含量的多少而定。然后加入蒸馏水多次水洗至中性[8]。
扫描电镜(SEM)观察: 吸取一定体积的酸化处理样品, 滴加在SEM铜台上, 自然晾干后, 溅射仪喷金, 然后即可在Philips XL-30 SEM下进行观察和拍照。
2 种类描述
本文观察到四种双唇形突海链藻, 分别是有翼海链藻T. bipartita (Rattray) Hellegraeff、脆弱海链藻T. fragilis Fryxell、亨氏海链藻T. hendeyi Hasle & Fryxell和维斯吉思海链藻T. visurgis Hustedt, 其中有三个为我国新记录种类: 脆弱海链藻、亨氏海链藻和维斯吉思海链藻。亨氏海链藻来自纯化的培养藻株, 其余来自自然样品。
2.1 有翼海链藻Thalassiosira bipartita (Rattray) Hallegraeff (图版Ⅰ)
Hallegraeff 1992, p.21, figs 1—12.
细胞单独生活。壳面圆形, 直径25—71 μm, 壳环带有零至数个胶质翼膜(图版Ⅰ-1、2)。壳面较平,壳套窄。孔纹直线状至偏心状排列(图版Ⅰ-3、6), 中央区域孔纹较大, 密度是10 μm大约4—5个; 边缘较小, 密度是10 μm内6—10个, 壳面中央有一个中央孔纹, 周围环绕着6个孔纹(图版Ⅰ-3、6)。壳面边缘有一圈显著的肋纹(图版Ⅰ-3—5), 高约3 μm,密度为10 μm内10—14个。两圈不规则排列的、外形较小的壳缘支持突位于肋纹上(图版Ⅰ-4、5), 壳缘支持突内壳面观排成一圈, 密度为10 μm内4—5个, 这些外形较小的壳缘支持突没有外管(图版Ⅰ-4, 5), 但具有短的内管(图版Ⅰ-8、9)。两个壳缘唇形突具有长且粗的外管(图版Ⅰ-4、5), 相距150°—180°。
生境: 海洋浮游生活。
采样点: 标本采自长江口海域(7月、8月)、广东珠海沿海(10月)、大亚湾(4月、12月)、香港海域(12月)。我国曾记录于福建东山到闽侯沿海, 海南岛附近海域[9]。本种为浅海暖水种, 分布较广, 在爪哇及沿海水域, 新加坡, 印度洋阿拉伯海, 泰国海湾, 卡奔塔利亚湾, 咸海, 可可群岛、日本南部等沿海水域均有记录[10,11]。
2.2 脆弱海链藻Thalassiosira fragilis Fryxell中国新记录(图版Ⅱ)
Fryxell et al. 1984, p.147, figs 17—28.
细胞环面观鼓形, 壳面圆形, 中央略隆起。壳面壁薄, 硅质化弱, 直径29—37 μm。壳面孔纹小, 束状排列(图版Ⅱ-10、11), 密度为10 μm 内40—50个。一个中央支持突, 细胞壳面二分之一处有4—11个支持突, 常见4个(图版Ⅱ-12、13)。一圈壳缘支持突, 密度10 μm 内3—4个, 所有支持突基部有4个围孔。两个壳缘唇形突(图版Ⅱ-12), 相距90°—170°。壳面所有支持突及唇形突均没有外管(图版Ⅱ-11), 但具有短的内管(图版Ⅱ-14、15)。有报道认为该种细胞可形成胶质块群体, 群体中细胞通过胶质丝相连[12]。但我们的样品由于细胞数量较少, LM下未观察到群体。
生境: 海洋浮游生活。
采样点: 标本采自浙江沿海水域(7月)。本种属于亚热带温水种, 首次记录于北大西洋[12]。
2.3 亨氏海链藻Thalassiosira hendeyi Hasle & Fryxell中国新记录(图版Ⅲ)
Hasle & Fryxell 1977, p.25, figs 35—45.
细胞单独生活, 或形成两个细胞的短链。细胞壳面圆形, 中央略凹(图版Ⅲ-16), 直径36—54 μm。环面观四边形, 贯壳轴小于直径, 环带较宽, 环带上的孔纹清晰可见(图版Ⅲ-16), 密度为10 μm大约有10个。壳面孔纹直线或略成偏心状排列(图版Ⅲ-18—20), 密度10 μm 内5—6个, 近壳缘处为10 μm内12—14个。通常有1个中央支持突(偶见2个或三个)位于筛孔之间的壁上(图版Ⅲ-18、21、23、25),有4—5个围孔, 支持突基部较高, 与其他海链藻不同(图版Ⅲ-21)。2圈排列不规则的壳缘支持突(图版Ⅲ-17、23、24), 密度10 μm 内5—6个, 基部有4个围孔。壳缘支持突具有短的外管(图版Ⅲ-22)和内管(图版Ⅲ-26、27)。壳面边缘或有肋纹, 密度为10 μm 内9—10个。两个壳缘唇形突, 偶见三个(图版Ⅲ-19), 具长的外管(图版Ⅲ-22), 唇形突与壳面中央支持突的连线夹角变化较大, 约90°—180°, 通常为150°—180°。
生境: 海洋浮游生活。
采样点: 标本采自香港水域(12月)。本种为温带亚热带广布种, 记录于非洲西部沿海[13], 墨西哥湾[14], 旧金山湾[15], 在北海附近海域[16]也有发现。
2.4 维斯吉思海链藻Thalassiosira visurgis Hustedt中国新记录(图版Ⅳ)
Hasle 1978, p.263, figs 1—4.
细胞直径为15—21 μm。壳面孔纹偏心状排列(图版Ⅳ-28、29), 在壳面中央处密度10 μm 内12—13个, 边缘处10 μm内约 15个。一个中央支持突(图版Ⅳ-28—30, 33—35), 基部有4个围孔(图版Ⅳ-35)。一圈壳缘支持突, 密度10 μm 内8—9个, 基部4个围孔, 壳缘支持突具有长的外管(图版Ⅳ-31、32), 但内管很短(图版Ⅳ-36、37)。两个唇形突具有长的外管(图版Ⅳ-31、32), 位于一圈壳缘支持突之间, 相距大约180°, 相对略远离靠缘一些。
生境: 海洋浮游生活
采样点: 样品采自香港海域(12月)及湛江海域(11月)。本种在淡水、海水中均有分布, 在德国[17],英格兰[18], 美国[15,19], 墨西哥湾[14]等地都有报道。
3 讨论
海链藻个体微小, 形态学区别细微, 准确鉴定的难度较大。主要的形态分类学依据有群体特征、细胞大小、孔纹排列方式, 以及壳面突起的类型、数量、密度和排列方式, 其中, 突起的形态学特征是关键性依据。突起有三类: 支持突、唇形突和闭合突。唇形突在海链藻中普遍存在, 绝大多数海链藻种类都只有一个唇形突, 双唇形突的现象仅见于极少数种类。目前为止, 国际上已报道的海链藻种类近400种[20], 而具有双唇形突的种类只有8种[6,11—13,19,21]。我国关于双唇形突海链藻的报道较少, 只有郭玉洁和钱树本记录了对称海链藻T. symmetrica Fryxell & Hasle[9], 但仅提供了双唇形突的光镜(LM)照片, 未能清晰显著两个唇形突的形态学特征。
对我国已记录的5个双唇形突海链藻的形态学特征进行了比较研究(表1)。双唇形突结构是本文记录种类的独有结构, 也是与相似海链藻种类的有效区别特征。除了双唇形突结构之外, 脆弱海链藻与细弱海链藻T. subtilis (Ostenfeld) Gran、双环海链藻T. diporocyclus Hasle相似: 壳面孔纹细密且束状排列、一环壳面支持突位于壳面中央至壳缘一半处,但脆弱海链藻的壳面硅质化相对更弱, 壳面一环支持突的数量较少, 常见4个, 而后两种的壳面支持突数量众多, 且不规则排列成1—2环。亨氏海链藻的壳面孔纹直线排列, 与海链藻直线亚属的种类相似, 如微小海链藻T. exigua Fryxell & Hasle、线形海链藻T. lineata Jousé、柔弱海链藻T. tenera Proschkinae-Lavrenko等, 双唇形突结构是有效区别特征。亨氏海链藻与同样具有双唇形突结构的T. simonsenii Hasle & Fryxell也较相似, 主要区别是前者的壳缘没有闭合突, 后者壳缘具有刺状闭合突。以往, 唇形突被认为是比较稳定的形态学特征, 而亨氏海链藻的唇形突则易变化, 主要体现在其数量和分布位置上。亨氏海链藻的唇形突数量通常为两个, 其与壳面中央支持突的连线夹角成150°—180°;而在一段时间的室内培养之后, 出现了具有三个唇形突的壳面(图版Ⅲ-19), 也有夹角成90°的(图版Ⅲ-19、24)。维斯吉思海链藻与众多典型海链藻种类相似, 即具有一个中央支持突、一环壳缘支持突的海链藻种类, 如艾伦海链藻T. allenii Takano、棱角海链藻T. angulata (Greg.) Hasle、太平洋海链藻T. pacifica Gran & Angst等种类, 双唇形突结构也是最为直观和有效的区别特征。
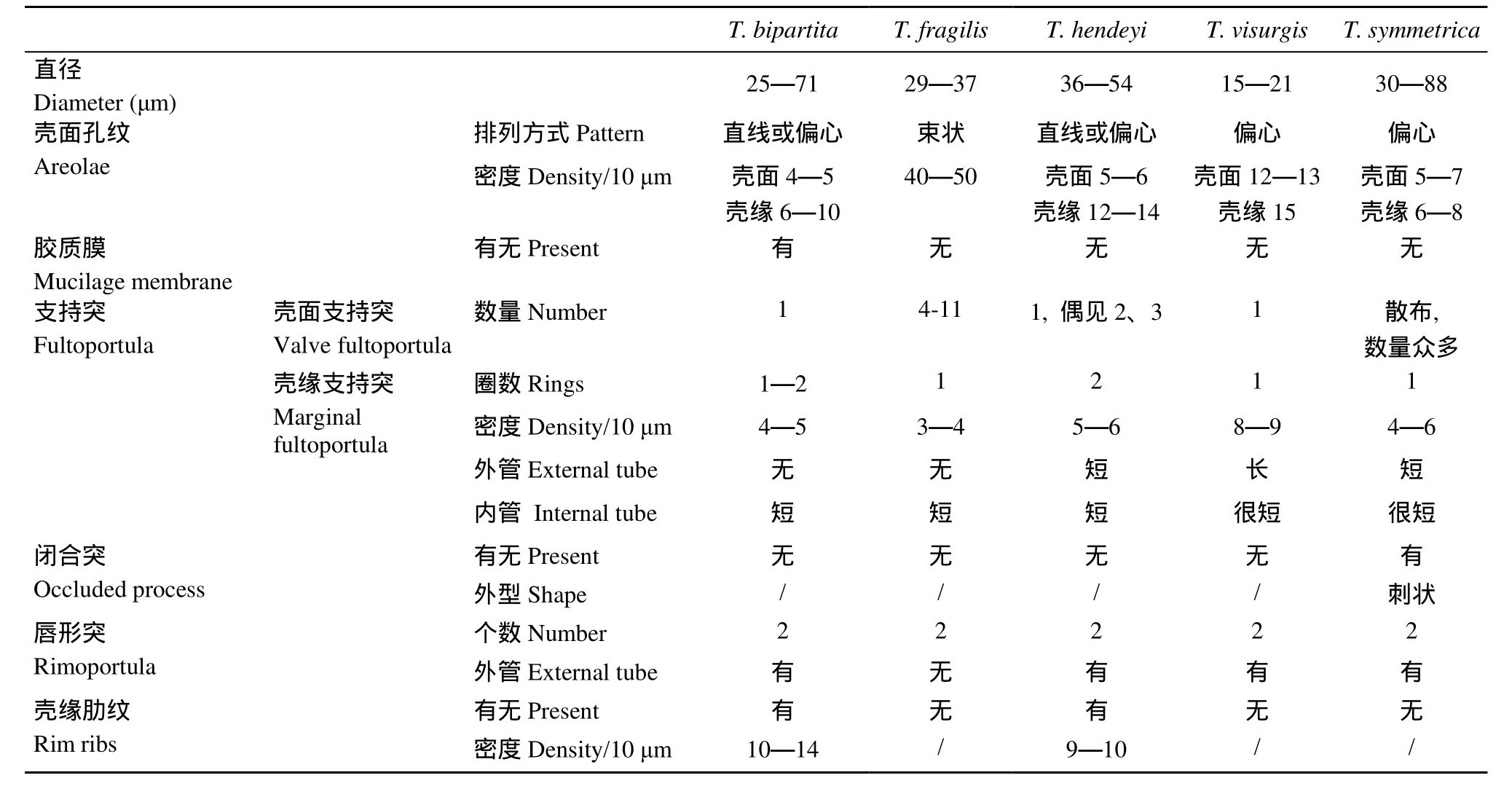
表1 我国已报道五种双唇形突海链藻的形态学比较Tab.1 Morphological comparison among five reported Thalassiosira species with two rimoportulae from China
与有翼海链藻具有相似壳面特征的种类不多,该种显著的壳缘肋纹结构, 以及位于壳环带的胶质膜状结构, 是本种的标志性区别特征。Hallegraeff将有翼圆筛藻修订为有翼海链藻的关键依据[11], 是认为本种具有数量众多的壳缘支持突, 而非数量众多的壳缘唇形突。金德祥等在我国多个海域记录了有翼圆筛藻Coscinodicus bipartitus Rattray[22], 由于并未记录标本是否具有双唇形突结构, 因此, 尚不能确定我国沿海分布的有翼圆筛藻是否为有翼海链藻。壳缘突起是以支持突为主, 还是唇形突为主, 是海链藻和圆筛藻之间的关键性区别特征。然而, 本种的修订带来另外一个问题, 以往认为海链藻与漂流藻Planktoniella的区别是壳环带胶质膜的有无,即漂流藻的标志性特征是具有显著的胶质膜, 有翼海链藻的修订则模糊了两属之间的这一区别, 并且漂流藻的壳面特征与有翼海链藻极为相似, 也具有数量众多的壳缘支持突和两个壳缘唇形突。Hallegraeff在修订了有翼海链藻之后, 重新界定了漂流藻种类与有翼海链藻的区别特征, 认为两者虽然都具有胶质膜[11], 但结构上不同, 有翼海链藻的胶质膜结构疏松, 而漂流藻胶质膜的结构致密, 且中间穿插有硅质肋。本文认为, 有翼海链藻的修订,弱化了胶质膜的分类学价值, 也模糊了海链藻与漂流藻的形态学界限, 目前还需要利用多种系统学研究手段, 寻找更多的依据, 以判断胶质膜是否具有稳定的分类学意义, 确认有翼海链藻的修订是否证据确凿。若胶质膜不具备稳定的分类学意义, 则需要依据同样的原则, 将整个漂流藻属修订到海链藻属之中, 这些都是后续工作的方向。
突起的数量和分布位置是海链藻形态分类学的关键依据, 并且已经被广泛应用到海链藻科, 甚至整个硅藻门的系统学研究[6]。唇形突在中心纲硅藻中极为常见, 其数量和分布位置相对稳定, 被认为是相对保守的结构, 赋予重要的分类学价值[7]。唇形突在壳缘的位置被认为是稳定的分类学依据, 应用于海链藻的形态分类, 如T. aestivalis Gran与T. concaviuscula Makarova之间唯一的区别特征就是唇形突的位置, 前者的唇形突位于两个壳缘支持突正中间, 后者则靠近其中一个壳缘支持突。然而对于其他海链藻种类, 唇形突位置存在一定程度的变化, 如诺氏海链藻T. nordenskioeldii Cleve的壳缘唇形突位置常有变化, 有时靠近一个壳缘支持突, 有时位于两个壳缘支持突正中间[23]。在以往报道中,亨氏海链藻的两个唇形突与壳面中央支持突的连线夹角成150°—180°, 本研究则观察到夹角成90°(图版Ⅲ-19, 24)。绝大部分海链藻的唇形突数量也是极为稳定的特征, 但也有少数变化的情况, 如双线海链藻的唇形突数量在1—3个[24]; 在本文的研究结果中, 亨氏海链藻的唇形突数量也有变化的情况,大多是两个, 偶有三个的。因此, 唇形突相关特征的变化情况及其分类学价值, 还需要其他研究手段的应用和验证。形态学与分子生物学技术相结合的研究手段已在拟菱形藻等硅藻类群中得到了广泛应用,并获得了显著的成果[25], 开展其他分类技术在海链藻中的应用, 可为唇形突分类学价值的判断提供重要的参考依据。此外, 唇形突数量的大尺度变化还是某些硅藻在属水平上的关键区别特征, 例如形态学上接近的海链藻属和圆筛藻属, 绝大部分海链藻种类只有1个唇形突, 极少数种类具有2个唇形突,而圆筛藻种类具有数量众多的唇形突, 唇形突的数量成为两个相似属种类的重要区分依据。双唇形突结构仅存在于少数海链藻种类, 而具有这一特殊特征的少数海链藻种类, 其系统学位置又如何?是否处于典型海链藻(单个唇形突)和圆筛藻(多个唇形突)之间?通过对双唇形突海链藻系统学的相关研究,可望阐述唇形突数目的分类学和系统学价值。
[1] Round F E, Crawford R M, Mann D G. The Diatoms: Biology and Morphology of the Genera [M]. Cambridge: Cambridge University Press. 1990, 747
[2] Fryxell G A, Hasle G R. Taxonomy of harmful diatoms [A]. In: Hallegraeff G M, Anderson D M, Cembella A D (Eds.), Manual on Harmful Marine Microalgae [M]. Paris: Imprimerie Landais (United Nations Educational, Scientific and Cultural Organization). 2004, 465—510
[3] Li Y. Taxonomic studies and ecological characteristics on Nano-diatoms in coastal waters of China [D]. Xiamen Uni-versity. 2006. [李扬. 中国近海海域微型硅藻的生态学特征和分类学研究. 厦门大学理学博士学位论文. 2006]
[4] Chen S W, Gao Y H, Du H, et al. First recording of Thalassiosira diporocyclus bloom in the southeast China sea [J]. Oceanologia et Limnologia Sinica, 2004, 35(2): 130—137 [陈善文, 高亚辉, 杜虹, 等. 双环海链藻(Thalassiosira diporocyclus Hasle)赤潮. 海洋与湖沼, 2004, 35(2): 130—137]
[5] Xie W L, Li Y, Gao Y H. First record of Thalassiosira curviseriata Takano (Bacillariophyta) and its bloom in the East China Sea [J]. Acta Oceanologica Sinica, 2008, 27(6): 124—132
[6] Hasle G R, Syvertsen E E. Marine Diatoms [A]. In: Tomas C R (Eds.), Identifying Marine Phytoplankton [M]. San Diego: Academic Press. 1997, 5—387
[7] Fryxell G A. Morphology, taxonomy and distribution of selected diatom species of Thalassiosira Cleve in the Gulf of Mexico and Antarctic waters [D]. Dissertation for the degree of Doctor of Philosophy, Texas A & M University. College Station, TX. 1975
[8] Cheng Z D, Gao Y H, Liu S C. Nano-diatoms in Fujian Coast [M]. Beijing: Ocean Press. 1993, 91 [程兆第, 高亚辉, 刘师成. 福建沿岸微型硅藻. 北京: 海洋出版社. 1993, 91]
[9] Guo Y J, Qian S B. Flora Algarum Marinarum Sinicarum, Vol.5, No.1 Centricae [M]. Beijing: Science Press. 2003, 493 [郭玉洁, 钱树本. 中国海藻志, Vol.5, No.1 中心纲. 北京:科学出版社. 2003, 493]
[10] Fukuyo Y, Takano H, Chihara M, et al. Red Tide Organisms in Japan—an Illustrated Taxonomic Guide [M]. Tokyo: Uchida Rokakuho, 1990, 407
[11] Hallegraeff G M. Observations on the mucilaginous diatom Thalassiosira bipartita (Rattray) comb. nov. from the tropical Indo-West Pacific [J]. Diatom Research, 1992, 7(1): 15—23
[12] Fryxell G A, Gould R W Jr, Watkins T P. Gelatinous colonies of the diatom Thalassiosira in Gulf Stream warm core rings, including T. fragilis, sp. nov. [J]. British Phycological Journal, 1984, 19(2): 141—156
[13] Hasle G R, Fryxell G A. The genus Thalassiosira, some species with a linear array [J]. Beiheft zur Nova Hedwigia, 1977, 54(1-2): 15—66
[14] Aké-Castillo J A, Hernández-Becerril D U, Meave Del C M E. Species of the Genus Thalassiosira (Bacillariophyceae) from the Gulf of Tehuantepec, Mexico [J]. Botanica Marina, 1999, 42(6): 487—503
[15] Mahood A D, Fryxell G A, McMillan M. The diatom genus Thalassiosira: species from the San Francisco Bay system [J]. Proceedings of the California Academy Sciences, 1986, 44(8): 127—156
[16] Hoppenrath M. Thalassiosira species (Bacillariophyceae, Thalassiosirales) in the North Sea at Helgoland (German Bight) and Sylt (North Frisian Wadden Sea), a first approach to assessing diversity [J]. European Journal of Phycology, 2007, 42(3):271—288
[17] Hustedt F. Die Diatomeenflora des Flusssystems der Weser im Gebiet der Hansestadt Bremen [J]. Abhadlungen herausgegeben vom Naturwissenschaftlichen Verein zu Bremen, 1957, 34: 181—440
[18] Belcher J H, Swale E M F. Notes on some small Thalassiosira (Bacillariophyceae) from the plankton of the lower Thames and other British estuaries (identified by transmission electron microscopy) [J]. European Journal of Phycology, 1986, 21(2): 139—145
[19] Hasle G R. Some freshwater and brackish water species of the diatom genus Thalassiosira Cleve [J]. Phycologia, 1978, 17(3): 263—292
[20] Catalogue of Diatom Names. (2009). California Academy of Sciences, On-line Version. Compiled by Elisabeth Fourtanier & J. Patrick Kociolek. Available online at http://www. calacademy.org/research/diatoms/names/index.asp
[21] Fryxell G A, Hasle G R. Thalassiosira eccentrica (Ehrenberg) Cleve, T. symmetrica sp. nov., and some related centric diatoms [J]. Journal of Phycology, 1972, 8(4): 297—317
[22] Chin D Q, Chen J H, Huang K G. Pelagic Diatoms in China Sea [M]. Shanghai: Shanghai Science and Technology Press. 1965, 230 [金德祥, 陈金环, 黄凯歌. 中国海洋浮游硅藻类. 上海: 上海科学技术出版社. 1965, 230]
[23] Hasle G R. Some Thalassiosira species with one central process (Bacillariophyceae) [J]. Norwegian Journal of Botany, 1978, 25(1): 77—110
[24] Pienaar C, Pieterse A J H. Thalassiosira duostra sp. nov. a new freshwater centric diatom from the Vaal River, South Africa [J]. Diatom Research, 1990, 5(1): 105—111
[25] Li Y, He L N, Ma Y Y, et al. The study on morphological taxonomy of Pseudo-nitzschia pseudodelicatissima complex [J]. Acta Hydrobiologica Sinica, 2010, 34(2): 302—311 [李扬, 何利娜, 马艳艳, 等. 伪柔弱拟菱形藻复合群的形态分类学研究. 水生生物学报, 2010, 34(2): 302—311]
MORPHOLOGICAL STUDIES OF THALASSIOSIRA SPECIES WITH TWO RIMOPORTULAE FROM CHINESE COASTAL WATERS
LI Yang1, ZHAO Qiu-Lan1,2and LÜ Song-Hui3
(1. Guangdong Provincial Key Laboratory of Healthy and Safe Aquaculture, Key Laboratory of Ecology and Environment Science in Guangdong Higher Education, College of Life Science, South China Normal University, Guangzhou 510631, China; 2. Department of Ceramic Technology, Hanshan Normal University, Chaozhou 521041, China; 3. Research Center for Harmful Algae and Aquatic Environment, College of Life Science and Technology, Jinan University, Guangzhou 510632, China)
Morphology and pattern of processes, including three types, fultoportula, rimoportula and occluded process, are key taxonomic characteristics for Thalassiosira. Rimoportula is a very common structure in all Thalassiosira taxa. Most Thalassiosira species only bear one rimoportula, and so far only 8 taxa have been reported to bear two rimoportulae from nearly 400 recorded Thalassiosira taxa. The quantity of rimoportulae has been considered as a different key morphological characteristic between the genus Thalassiosira and Coscinodiscus. Previous reports about Thalassiosira species with two rimoportulae from Chinese coast were rare, and almost no electron microscopy photos about the type of two rimoportulae are available. In order to characterize the species diversity and distribution of the genus Thalassiosira along Chinese coastal waters, some water samples were collected from several representative locations. Thalassiosira specimens within these samples, as well as several clonal strains isolated from natural samples, were studied by scanning electron microscopy. Four Thalassiosira species with two rimoportulae were observed and they were T. bipartita (Rattray) Hallegraeff, T. fragilis Fryxell, T. hendeyi Hasle & Fryxell and T. visurgis Hustedt. Three of them were newly recorded species for China, T. fragilis, T. hendeyi and T. visurgis. The detailed morphological description, habits and ecological distribution of these species were given in this paper. Furthermore, taxonomic comparison among resembling species were made. The transfer of T. bipartita from C. bipartita obscured the morphological difference between the genus Thalassiosira and Planktoniella, and the taxonomic estimation of mucilage membrane should be re-examined in the future. Some morphological variations of marginal rimoportulae and central fultoportulae in T. hendeyi were observed, and analogous variations in other Thalassiosira species were discussed. The taxonomic evaluations of rimoportulae, including their quantities and locations, should be re-analyzed in conjunction with the usage of other modern techniques.
Thalassiosira; Two rimoportulae; Morphology; Chinese coastal waters

图1—5为外壳面观; 图1—2为具胶质翼状膜的壳面; 图3为翼状膜脱落的壳面, 示两个唇形突和一个中央支持突的位置; 图4—5示唇形突及其外管, 以及位于壳缘肋纹上的支持突; 图6—9为内壳面观; 图6示两个唇形突和一个中央支持突的位置; 图7示中央支持突具有5个围孔; 图8—9唇形突及其内管, 以及壳缘支持突及其内管. 图中窄箭头示支持突, 宽箭头示唇形突. 比例尺标于图内Figs 1—5. Outside valve views; Figs 1—2. Whole valves with mucilage membranes; Fig. 3. Whole valve lacking mucilage membrane, showing the locations of two rimoportulae and one central fultoportula; Figs 4—5. Rimoportulae with long external tubes and marginal fultoportulae on the ribs; Figs 6—9. Inside valve views; Fig.6. Locations of two rimoportulae and one central fultoportula; Fig.7. One central fultoportula with five satellite pores; Figs 8—9. Rimoportulae with internal slits and marginal fultoportulae with internal tubes. Narrow arrow means fultoportula. Broad arrow means rimoportula. Scale bar is shown in each figure
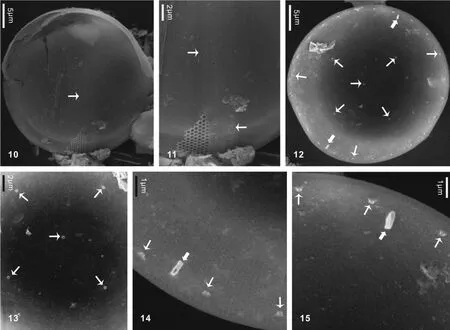
图10—11为外壳面观; 图10示整个外壳面; 图11示孔纹排列及壳面近中央处的支持突; 图12—15为内壳面观; 图12示两个唇形突和支持突的分布; 图13示一个中央支持突和四个壳面支持突; 图14—15示唇形突和壳缘支持突及其内管. 图中窄箭头示支持突, 宽箭头示唇形突. 比例尺标于图内Figs 10—11. Outside valve views. Fig.10. Whole outside valve; Fig.11. Fasciculate pattern of areolae and fultoportulae near valve center; Figs12—15. Inside valve views; Fig.12. Locations of two rimiportulae and fultoportulae in the internal valve; Fig.13. One central fultoportula and four sub-central fultoportulae; Figs14—15. Rimoportulae with internal slits and marginal fultoportulae with internal tubes. Narrow arrow means fultoportula. Broad arrow means rimoportula. Scale bar is shown in each figure

图16、图17为壳环面观; 图16示唇形突的侧面观; 图17示不规则分布的壳缘支持突; 图18—22为外壳面观; 图18示两个唇形突和一个中央支持突的分布; 图19示三个唇形突和一个中央支持突的分布; 图20示两个唇形突和三个中央支持突的分布; 图21示三个中央支持突; 图22示唇形突和支持突及其外管; 图23—27为内壳面观; 图23—24示唇形突和支持突在内壳面上的分布; 图25示两个中央支持突; 图26—27示唇形突内裂缝, 支持突及其内管. 图中窄箭头示支持突, 宽箭头示唇形突. 比例尺标于图内Figs 16, 17. Girdle views; Fig.16. Lateral view of rimoportulae with long external tubes; Fig.17. Marginal fultoportulae arranged into irregular ring; Figs 18—22. Outside valve views; Fig.18. Valve with two rimoportulae and one central fultoportula; Fg.19. Valve with three rimoportulae and one central fultoportula; Fig.20. Valve with two rimoportulae and three central fultoportulae; Fig.21. Enlargement of three central fultoportulae; Fig.22. Rimoportula and fultoportulae with external tubes; Figs 23—27. Inside valve views; Figs 23—24. Locations of rimoportulae and fultoportulae; Fig.25. Enlargement of two central fultoportulae; Figs 26—27. Rimoportulae with internal slits and fultoportulae with internal tubes. Narrow arrow means fultoportula. Broad arrow means rimoportula. Scale bar is shown in each figure

图28—32为外壳面观; 图28—29示唇形突和支持突的分布; 图30示一个中央支持突; 图31—32示唇形突和支持突的外管; 图33—37为内壳面观; 图33—34示唇形突和支持突的分布; 图35示一个中央支持突及其四个围孔; 图36—37示唇形突和支持突的内管. 图中窄箭头示支持突, 宽箭头示唇形突. 比例尺标于图内Figs 28—32. Outside valve views; Figs 28—29. Locations of rimoportulae and fultoportulae on external valves; Fig.30. One central fultoportula; Figs 31—32. Rimoportulae and fultoportulae with external tubes; Figs 33—37. Inside valve views; Figs 33—34. Locations of rimoportulae and fultoportulae on internal valves; Fig.35. One central fultoportula with four satellite pores; Figs 36—37. Rimoportulae with slits and marginal fultoportulae with internal tubes. Narrow arrow means fultoportula. Broad arrow means rimoportula. Scale bar is shown in each figure
Q94
A
1000-3207(2013)01-0106-10
10.7541/2013.106
2011-09-02;
2012-10-17
国家自然科学基金(30700046); 广东省自然科学基金(S2012010008456)资助
李扬(1978—), 男, 河南信阳人; 副研究员, 博士; 主要从事藻类分类学和生态学研究。E-mail: liyang@scnu.edu.cn
吕颂辉, E-mail: lusonghui1963@163.com
