海拔对钝裂银莲花不同花色居群间繁殖特征及繁殖分配的影响
2013-04-12李冰刘左军赵志刚胡春任红梅伍国强
李冰,刘左军*,赵志刚,胡春,任红梅,伍国强
(1.兰州理工大学生命科学与工程学院,甘肃 兰 州730050;2.兰州大学干旱与草地生态教育部重点实验室,甘肃 兰 州730000)
植物的繁殖对策是指植物在其生活史过程中,通过最佳的资源分配格局,以其特有的繁殖属性去适应环境,提高植物适合度的自组织过程[1]。植物的繁殖性状同时受到内在遗传因子和外在环境因子的影响。而环境因子随海拔梯度发生的剧烈变化为研究植物的生长和繁殖的生态适应性提供了理想的条件[2]。如随着海拔的升高,环境温度、生长季长度和资源有效性均会减少,而年降水量、霜冻发生频率及太阳辐射量则会增加[3],昆虫的丰富度、多样性以及活动性会降低[4],可能导致高海拔虫媒植物受花粉限制[5]。因此,植物的繁殖策略也会随之发生改变。根据最优分配模型理论分析,多年生植物的繁殖分配应随海拔的升高而减小[6-8]。但多数研究又发现[9-12],植物的繁殖分配随海拔的升高而增大,显示有性繁殖具有更为重要的作用[12-15]。当然,也有研究表明,植物繁殖分配模式随海拔升高并不总是呈现出一致性的变化规律[16]。
很多虫媒植物都有不同的花色性状,花色对传粉者具有吸引和引导作用[17,18]。传粉昆虫对花色的反应和嗜好与植物发展和繁荣密切相关[19]。如翠雀属Delphiniumnelsonii和番薯属Ipomoeapurpurea居群中,传粉昆虫表现出负频率依赖性选择[18,20]。但在兰科植物Caladeniabehrii居群中,并不存在这种负频率依赖性选择[21]。
植物的生长具有可塑性[22],其在自然种群中总表现出大小等级性。而植物个体大小一般也会随着海拔的升高而减小[23]。大量研究表明[8-12,16,24],同一物种繁殖性状在不同海拔间均存在大小依赖性现象,但其影响机制不尽相同。因此,在研究海拔对植物繁殖特征和繁殖分配的大小依赖性影响时,需要探讨该影响是通过个体大小间接产生,还是直接作用。
综上所述,国内外有关海拔对植物繁殖特征及繁殖分配模式的影响主要集中在不同物种间的比较,以及植物种内不同花色的地理分布和花色对传粉者是否具有吸引作用等方面。如果花色对传粉者有吸引作用,说明不同花色居群间的繁殖特征及繁殖分配可能存在差异。但有关植物种内不同花色居群间是否具有相同的繁殖对策、以及对海拔的变化有何响应等方面的研究尚未见报道。
钝裂银莲花(Anemoneobtusiloba)主要分布于西藏南部和东部、四川西部,在尼泊尔、锡金、不丹、印度北部也有分布[25],是高寒草甸中常见的一种毒杂草,花色从白色到黄色差异很大。目前,国内外对钝裂银莲花的研究主要集中在不同环境下,同属不同物种间繁殖分配和性分配的比较[8,26-28]。选择钝裂银莲花不同花色居群为研究对象,探讨海拔对其繁殖特征、繁殖分配的影响,以及当繁殖压力增大时又会采取什么样的策略来达到繁殖目的,为高寒草甸植物生活史对策研究和防治草地退化提供理论依据。
1 材料与方法
1.1 研究区域概况
研究区在甘肃省甘南藏族自治州,位于青藏高原东部地区(101°~103°E,34°~35°70′N),海拔2 900~4 000 m;年均降水量450~780 mm,降水主要集中在7-9月;年平均气温为1.8℃,1月份平均温度-10.7℃,7月份平均温度11.7℃,生长季最高气温23.6~28.9℃;年平均霜期不少于270 d。草地类型以高寒草甸为主,该区地势开阔、多风、气候寒冷等[7,29,30]。
1.2 材料
试验材料为青藏高原东部高寒草甸区分布的常见毛茛科植物钝裂银莲花。钝裂银莲花是毛茛科(Ranunculaceae)毛茛亚科银莲花属(Anemone)虫媒两性花多年生植物。花白色、浅黄色或黄色(图1),心皮和雄蕊多数。花期在6-7月,果期7-8月;花期一般1周左右,果期在2周以上。
1.3 取样和测量
2011年6月在甘南境内沿海拔梯度选取4个样区(表1)。在每个样区分别随机采集白色、浅黄色和黄色钝裂银莲花盛花期植株各40~50株,取其地上部分。所有植株均在自然生境下收集。

图1 不同颜色的钝裂银莲花Fig.1 Different colors of A.obtusiloba
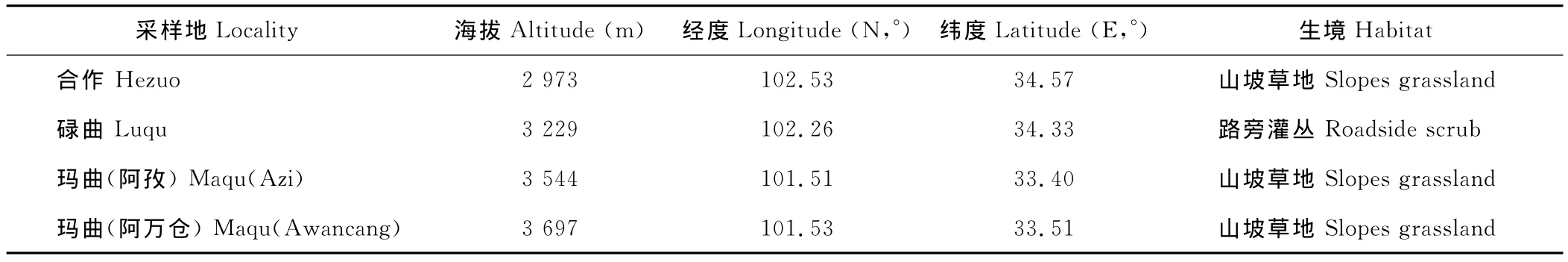
表1 钝裂银莲花居群概况Table 1 The background of field populations studied of A.obtusiloba
对于采集的样品,统计花期的总花数,测定单花面积、花展示面积。总花数为所有花芽、开放和凋萎的花数目。每个植株上正在盛开的花数目,以n表示;植株上每个盛开的花面积用A表示;单花面积用表示。植株上每个盛开的花面积用公式A=d1×d2计算[9],式中,d1为对称轴上的最大距离;d2为与该对称轴垂直方向上的最大距离。对同一植株上所有盛开的花面积进行测量,取其平均值代表该植株的单花面积,花展示面积就为×n。全部样品在80℃下烘烤24 h后,分开称量花、茎和叶的干重。
1.4 数据分析
植物的地上生物量以营养体(茎、叶)干重与总花干重的总和表示;个体大小以地上营养体(茎、叶)干重表示;花、茎、叶分配分别以总花干重、茎干重和叶干重占地上生物量的百分比表示;繁殖分配以总花干重占地上营养部分干重的百分比表示。用Bivariate correlations和Linear regression分析繁殖特征、繁殖分配与海拔的相关性和线性回归。在对不同海拔间繁殖特征和繁殖分配进行分析时,根据个体大小和上述繁殖组分的异速关系,用异速模型Y=a Xb分析个体大小(X)与繁殖组分(Y)的相关性[31,32]。Y=a Xb进行对数转化后得到线性方程:log10Y=log10a+blog10X,式中,b代表斜率;log10a为截距。进行数据分析时对个体大小、繁殖特征及繁殖分配的数据均进行了对数转换。用Liner regression进行线性回归分析,对于繁殖特征、繁殖分配与个体大小在海拔间异速关系都显著(P<0.05)的,本研究以个体大小为协变量,用General lineal model菜单中的Univariate比较不同海拔种群间斜率b和截距log10a的差异:如果截距log10a在海拔间有显著变化(P<0.05),则表明显著的海拔效应;如果斜率b在海拔间有显著变化(P<0.05),则表明海拔和个体大小的交互作用显著,说明繁殖特征或繁殖分配的大小依赖性随海拔的增高有显著的变化[26]。统计分析均采用SPSS 19.0统计软件。
2 结果与分析
2.1 海拔与地上生物量分配的关系
随着海拔的升高,3种花色钝裂银莲花地上生物量分配到花上的部分所占的比例均升高(图2),其中白色和黄色钝裂银莲花分别达到显著(P<0.05)和极显著水平(P<0.01),浅黄色未达到显著(P>0.05);分配到茎上部分所占比例随海拔的升高也都呈正相关,其中白色达到极显著水平(P<0.01),浅黄色和黄色钝裂银莲花均未达到显著水平(P>0.05)(图3);3种花色钝裂银莲花分配到叶上的部分所占比例却均随着海拔的升高呈负相关,其中白色达到极显著(P<0.01),浅黄色和黄色均未达到显著水平(P>0.05)(图4)。因此,随着海拔的升高,繁殖部分所占地上生物量的比例增大,说明植物对繁殖部分的投入也越大。
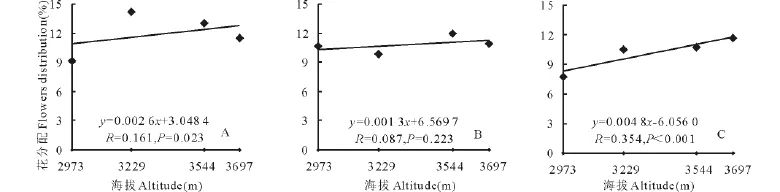
图2 海拔与花分配回归分析Fig.2 Relationships between flowers distribution and altitude
2.2 海拔与繁殖特征及繁殖分配的关系
随着海拔的升高,3种花色钝裂银莲花花数目均减少,浅黄色和黄色钝裂银莲花均达到极显著水平(P<0.01),而白色钝裂银莲花未达到显著水平(P>0.05)(图5);3种花色钝裂银莲花的单花面积(图5)和单花重(图6)均随着海拔的升高而极显著增大(P<0.01);海拔与3种花色的花展示面积(图6)、繁殖投入(图7)均呈正相关,但都未达到显著水平(P>0.05);3种花色的繁殖分配随海拔的升高呈正相关,其中白色和黄色钝裂银莲花分别达到显著(P<0.05)和极显著水平(P<0.01),而浅黄色未达到显著(P>0.05)水平(图7)。
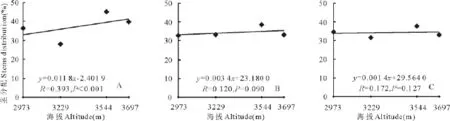
图3 海拔与茎分配回归分析Fig.3 Relationships between stems distribution and altitude
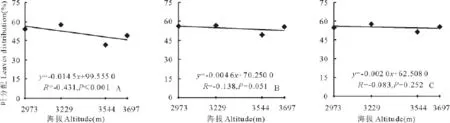
图4 海拔与叶分配回归分析Fig.4 Relationships between leaves distribution and altitude
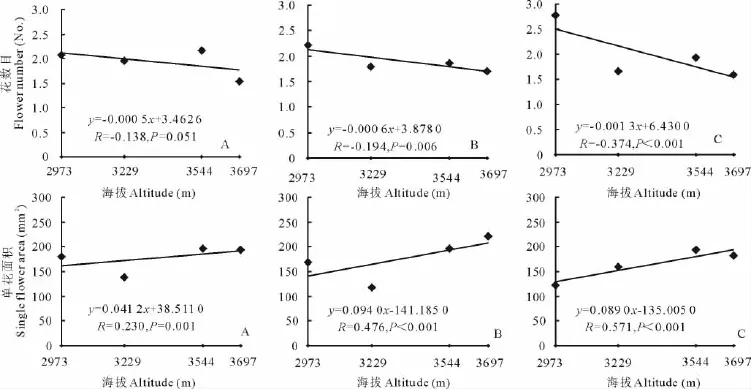
图5 海拔与不同花色钝裂银莲花花数目、单花面积的回归分析Fig.5 Relationships between flower number and single flower area of different colors A.obtusiloba and altitude
2.3 个体大小与繁殖特征及繁殖分配的关系
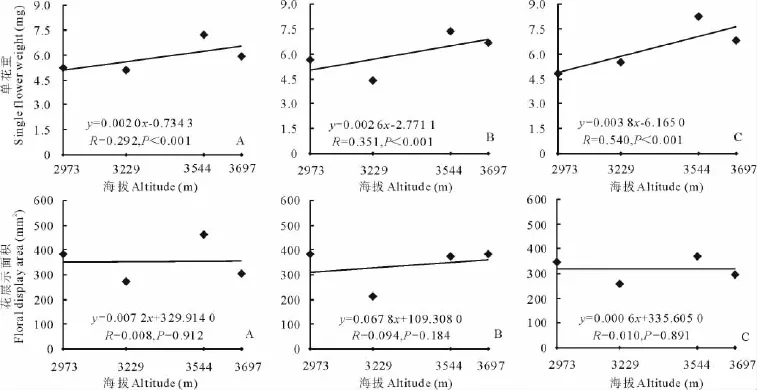
图6 海拔与不同花色钝裂银莲花单花重、花展示面积的回归分析Fig.6 Relationships between single flower weight and floral display area of different colors A.obtusiloba and altitude
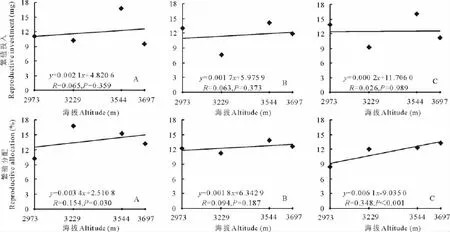
图7 海拔与不同花色钝裂银莲花繁殖分配的回归分析Fig.7 Relationships between reproductive allocation of different colors A.obtusiloba and altitude
分析结果显示(表2),在不同海拔下,3种花色钝裂银莲花花数目、单花重、花展示面积、繁殖投入与个体大小都表现出极显著正相关(P<0.01),单花面积与个体大小表现出显著正相关(P<0.05),且上述繁殖特征大小依赖的斜率在海拔间的变化不显著(P>0.05),但截距都有极显著(P<0.01)差异(除浅黄色钝裂银莲花繁殖投入在海拔3 697 m处表现为显著)。这说明3种花色钝裂银莲花花数目、单花面积、单花重、花展示面积、繁殖投入是大小依赖的,这些个体大小依赖的繁殖特征在海拔间的差异是海拔直接作用的结果,而不是海拔影响个体大小产生的。
除浅黄色钝裂银莲花在海拔3 697 m处繁殖分配与个体大小达到显著的负相关(P<0.05)外,3种花色钝裂银莲花在不同海拔下的繁殖分配与个体大小的关系均不显著(P>0.05)。可见,对于钝裂银莲花,虽然花期繁殖体重量确实随植株个体的增大而极显著增大,但是繁殖分配与个体大小间则没有表现出固定的相关模式。
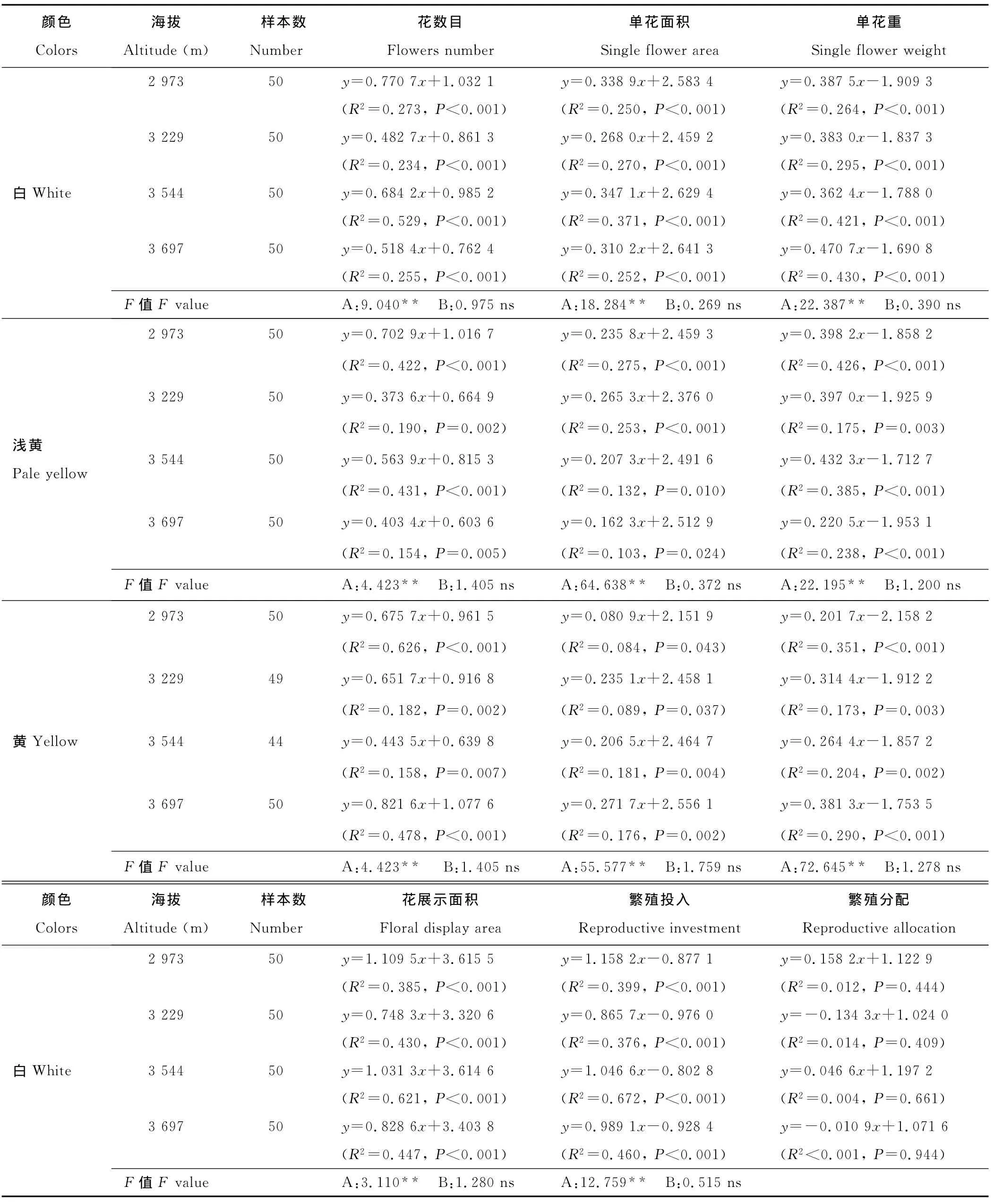
表2 不同海拔种群内个体大小与繁殖特征及资源分配的回归Table 2 Regressions of plant size and floral traits and resource allocation within different populations among altitudes
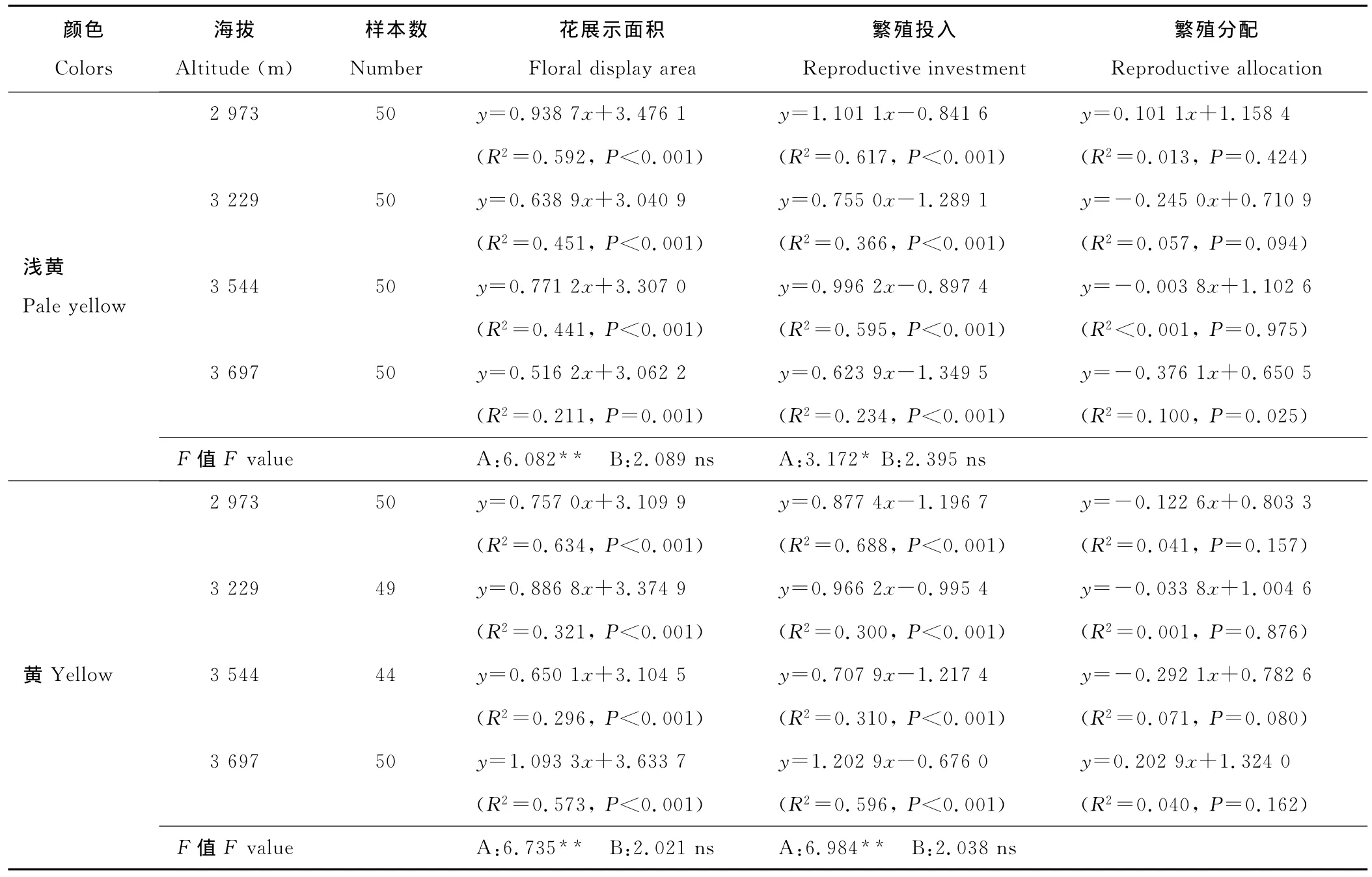
续表2 Continued
3 讨论
3.1 海拔与地上生物量分配的关系
海拔越高,不同花色钝裂银莲花对花和茎的投入均越大,对叶的投入越小。说明了不同花色钝裂银莲花资源分配对高海拔的恶劣环境做出了相同的适应性调整。在低海拔地区,茎的主要作用是支撑叶捕捉光,从而进行光合作用。而在高海拔地区,茎的主要作用是支撑花[13]。由于随着海拔的升高,植物生长期缩短,植物为了完成生活史,从而尽可能地减少营养结构的投入,而增加繁殖结构的投入。因此,植物只有增大对茎的投入才能支撑不断增大的繁殖结构。
3.2 海拔与繁殖特征及繁殖分配的关系
3种花色钝裂银莲花的花数目虽然随着海拔升高均在减少,但单花面积极显著增加,花展示面积有所增加,说明了高海拔地区植物对其吸引结构的投入增加,这与赵志刚[27]的结论相符。3种花色钝裂银莲花的单花重和繁殖投入均随着海拔的升高而增大,这与苏梅等[9]、Kud和 Molau[5]的研究结果相符,但不同于樊宝丽等[8]、Totland[33]的研究结果。3种花色钝裂银莲花繁殖分配均随着海拔的升高而增大,这与某些多年生植物的繁殖分配随海拔的升高而增加一致[10-11],却与樊宝丽等[8]的钝裂银莲花的繁殖分配随海拔的升高而减小的结论不同。
对于钝裂银莲花繁殖特征及繁殖分配随海拔的变化与樊宝丽等[8]的研究结果不同,可能存在以下原因:1)土壤湿度不同[23]。由于采样样地植被的覆盖度可能不同,使得土壤地表湿度不同,从而导致了繁殖器官的相对资源量和繁殖特征不同[34]。2)土壤养分不同。土壤因子与植物的表型性状的变异密切相关[35],高海拔地区可能有更高的土壤养分[26],有足够的矿物质资源分配给繁殖部分[13],从而对3种花色钝裂银莲花繁殖特征和繁殖分配造成了影响。3)传粉环境可能不同。随着海拔的升高,虽然面对不利的传粉环境且可利用的总资源量有限,保障有性繁殖的资源投入,才能保障种群在恶劣环境中的遗传变异性和进化适应潜力[13-15]。4)由于植物的可塑性,可能受到了植物自身遗传的影响,这些可塑性响应影响着植物的适合度,从而影响杂草的生存和繁衍[36]。5)对于多年生草本植物,地下部分储存的资源量对植株地上部分构件之间的资源分配也存在较为复杂的影响[16]。所以,海拔并不是影响植物繁殖分配策略的唯一作用因子,不同的生境环境和植物的遗传特性都与繁殖策略密切相关。因此,需要对不同海拔居群物种类型、种群密度、土壤水分和养分进行深入的分析,并结合交互移栽、传粉者访问频率、人工控制实验等,才能更全面的解释繁殖分配与遗传因素和生境环境间的关系。
3.3 个体大小与繁殖特征及繁殖分配的关系
在不同海拔梯度下,钝裂银莲花个体大小与3种花色花数目、单花重、花展示面积和繁殖投入均呈极显著正相关,与单花面积均呈显著正相关。这与他人研究[8,9]的结论相符。在多花植物中花大小通常随着植物个体的增大而增大[37,38]。植物在一个生长季中生产的花数目通常也随植物个体增大而增加[37,39,40]。花数目越多,说明了产生后代繁衍的可能性就越大;单花面积和花展示面积越大,说明增大了对传粉者的吸引,进而能够提高花粉的利用率及交配系统的多样性;单花重和繁殖投入越多,说明大个体植物将更多的资源投入到了繁殖器官,这些均表现出了植物的繁殖适合度的提高。海拔对3种花色钝裂银莲花繁殖特征的个体大小依赖性具有直接的影响,说明了各海拔生境中植物个体大小与繁殖特征的关系只是某些环境因素造成的,并不是自然选择的结果,因此不具有遗传基础。
大量的研究发现,植物的繁殖分配与个体大小之间可能存在负相关[7-10,28,41]、正相关[42]或无显著相关性[30]3种关系。在不同海拔下,对于不同花色钝裂银莲花而言虽然花期繁殖投入确实随植株个体大小的增大而极显著增加,但繁殖分配与个体大小则没有表现出固定的相关模式,表明不同颜色钝裂银莲花在不同海拔间并不存在大小依赖的繁殖分配。首先,这可能是由于不同海拔生境环境条件的不同导致了植物对确保生存的营养器官投入与繁殖器官投入进行了适应性调整,即植物根据自身的可塑性通过对繁殖分配策略做出了适应性调整来最大限度的保障其繁殖成功。其次,植物所处的环境条件恶劣,其生存和繁殖会受到外界因素的干扰,如牛羊的践踏、冰雹的损伤等[28]。最后,这3种花色性状不同的银莲花均属于同一物种,因此,繁殖分配还会受到系统发育水平等遗传方面的影响。不同花色钝裂银莲花繁殖分配对策是否与其所处的系统进化水平有关,且是否呈一定的规律性,还需进行更多的后续研究。
4 结论
海拔对不同花色钝裂银莲花的繁殖特征和繁殖分配有着重要的影响,且不同花色钝裂银莲花繁殖特征和繁殖分配对海拔的升高均做出了提高繁殖适合度的适应性调整,但海拔并不是影响钝裂银莲花繁殖策略的唯一因子,可能会受到环境、土壤、个体及物种间竞争和遗传等方面的影响。
致谢:在本实验过程中,兰州大学高寒草甸与湿地生态系统定位研究站为实验的正常展开提供了保障条件。在此表示衷心的感谢!
[1] 李金花,潘浩文,王刚.草地植物种群繁殖对策研究[J].西北植物学报,2004,24(2):352-355.
[2] Billings W D.Adaptations and origins of alpine plants[J].Arctic and Alpine Research,1974,6(2):129-142.
[4] Bingham R A,Ortnner A R.Efficient pollination of alpine plants[J].Nature,1998,391:238-239.
[6] Iwasa Y,Cohen D.Optimal growth schedule of a perennial plant[J].The American Naturalist,1989,133(4):480-505.
[7] 梁艳,张小翠,陈学林.多年生龙胆属植物个体大小与花期资源分配研究[J].西北植物学报,2008,28(12):2400-2407.
[8] 樊宝丽,孟金柳,赵志刚,等.海拔对青藏高原东部毛茛科植物繁殖特征和资源分配的影响[J].西北植物学报,2008,28(4):805-811.
[9] 苏梅,齐威,阳敏,等.青藏高原东部大通翠雀花的花特征和繁殖分配的海拔差异[J].兰州大学学报(自然科学版),2009,45(2):61-65.
[10] 张林静,石云霞,潘晓玲.草本植物繁殖分配与海拔高度的相关分析[J].西北大学学报(自然科学版),2007,37(1):77-80.
[11] 赵方,杨永平.中华山蓼不同海拔居群的繁殖分配研究[J].植物分类学报,2008,46(6):830-835.
[12] 孟丽华,王政昆,刘春燕,等.高山植物圆穗蓼的繁殖资源分配[J].西北植物学报,2011,31(6):1157-1163.
[13] Fabbro T,Krner C.Altitudinal differences in flower traits and reproductive allocation[J].Flora,2004,199:70-81.
[14] 范邓妹,杨永平.不同生境下珠芽蓼(蓼科)的繁殖策略比较[J].云南植物研究,2009,31(2):153-157.
[15] Fan D M,Yang Y P.Altitudinal variations in flower and bulbil production of an alpine perennial,Polygonumviviparum(Polygonaceae)[J].Plant Biology,2009,11(3):493-497.
[16] 王赟,胡莉娟,段元文,等.岩白菜(虎耳草科)不同海拔居群的繁殖分配[J].云南植物研究,2010,32(3):270-280.
[17] Smithson,Macnair M.Frequency-dependent selection by pollinators:mechanism and consequences with regard to behaviour of bumblebeesBombusterrestris(L.)(Hymenoptera:Apidae)[J].Journal of Evolutionary Biology,1996,9:571-588.
[18] Waser N M,Price M V.Pollinator choice and stabilizing selection for flower color inDelphiniumnelsonii[J].Evolution,1981,35:376-390.
[19] 钦俊德.昆虫与植物的关系-论昆虫与植物的相互作用及其演化[M].北京:科学出版社,1987.
[20] Brown B A,Clegg M T.Influence of flower color polymorphism on genetic transmission in a natural population of the common morning glory,Ipomoeapurpurea[J].Evolution,1984,38:796-803.
[21] Dickson C R,Petit S.Effect of individual height and labellum colour on the pollination ofCaladenia(syn.Arachnorchis)behrii(Orchidaceae)in the northern Adelaide region,South Australia[J].Plant Systematics and Evolution,2006,262:65-74.
[22] Happer J L.The Population Biology of Plants[M].San Diego:Academic Press,1977:424-449.
[23] Marcos M,Traveset A.Sexual allocation in single-flowered hermaphroditic individual in relation to plant and flower size[J].Oecologia,2003,137:69-75.
[24] 廖万金,张国全,张大勇.不同海拔藜芦种群繁殖特征的初步研究[J].植物生态学报,2003,27(2):240-248.
[25] 中国科学院中国植物志编辑委员会.中国植物志[M].北京:科学出版社,1980:34-36.
[26] Zhao Z G,Du G Z,Zhou X H,etal.Variations with altitude in reproductive traits and resource allocation of three Tibetan species of Ranunculaceae[J].Australian Journal of Botany,2006,54:691-700.
[27] 赵志刚.青藏高原高寒草甸常见毛茛科植物繁殖对策研究[D].兰州:兰州大学,2006:5.
[28] 赵志刚,杜国祯,任青吉.5种毛茛科植物个体大小依赖的繁殖分配和性分配[J].植物生态学报,2004,28(1):9-16.
[29] 陈学林,景国海,郭辉.青藏高原东缘高寒草甸19种马先蒿植物皮纹饰特征及其生物学意义[J].草业学报,2007,16(2):60-68.
[30] 陈学林,梁艳,齐威,等.一年生龙胆属植物的繁殖分配及其花大小、数量的权衡关系研究[J].草业学报,2009,18(5):58-66.
[31] Klinkhamer P G L,De Jong T J,Metz H.Sex and size in cosexual plants[J].Trends in Ecology and Evolution,1997,12:260-265.
[32] Wright S I,Barrett S C H.Size-dependent gender modification in a hermaphroditic perennial herb[J].Proceedings of the Royal Society Ser B,1999,266:225-232.
[33] Totland O.Environment-dependent pollen limitation and selection on floral traits in an alpine species[J].Ecology,2001,82(8):2233-2244.
[34] 黄德青,于兰,张耀生,等.祁连山北坡天然草地地上生物量及其与土壤水分关系的比较研究[J].草业学报,2011,20(3):20-27.
[35] 张妙青,王彦荣,张吉宇,等.垂穗披碱草种质资源繁殖相关特性遗传多样性研究[J].草业学报,2011,20(3):182-191.
[36] 王斌世,张荣.半干旱区农田杂草的生活史对策研究[J].草业学报,2011,20(1):257-260.
[37] Worley A C,Baker A M,Thompson J D,etal.Floral display inNarcissus:Variation in flower size and number at the species,population,and individual levels[J].International Journal of Plant Sciences,2000,161:69-79.
[38] Worley A C,Berrett S C H.Evolution of floral display inEichhorniapaniculata(Pontederiaceae):Genetic correlations between flower size and number[J].Journal of Evolutionary Biology,2001,14:469-481.
[39] Worley A C,Berrett S C H.Evolution of floral display inEichhorniapaniculata(Pontederiaceae):Direct and correlated response to selection on flower size and number[J].Evolution,2000,54:1533-1545.
[40] Sato H,Yahara.Trade-offs between flower number and investment to a flower in selfing and outcrossing varieties ofImpatienshypophylla(Balsalminaceae)[J].American Journal of Botany,1999,86:1699-1707.
[41] 刘左军,杜国祯,陈家宽.黄帚橐吾花序结构的资源配置与环境的关系[J].植物生态学报,2003,27(3):344-351.
[42] Hautier Y,Randin C F,Stocklin J,etal.Changes in reproductive investment with altitude in an alpine plant[J].Journal of Plant Ecology,2009,2(3):125-134.
