不同土地利用方式对典型温性草原群落物种组成和多样性以及生产力的影响
2013-04-12李愈哲樊江文张良侠翟俊刘革非李佳
李愈哲,樊江文,张良侠,翟俊,刘革非,李佳
(1.中国科学院地理科学与资源研究所,北京100101;2.中国科学院研究生院,北京100049)
草地是世界最广布的植被类型之一,面积约占全球陆地总面积的1/5[1]。中国的草原面积为3.55×108hm2,约占国土面积的41.7%,是世界草地总面积的6%~8%,居世界第二[2,3]。了解土地利用变化对草原群落物种组成,生态系统物种多样性、生产力有助于厘清人类活动对自然群落的改变,对于指导和改善人类对草原的利用、开发具有重要意义。锡林郭勒草原位于内蒙古自治区中部,地带性植被类型为典型温性草原,该草原类型约占到内蒙古天然草地总面积的1/3[4]。草原地区是自然环境严苛、生态系统稳定性相对脆弱的典型区域之一,不合理的人为活动已使该区50%左右的草原处于不同程度的退化之中[5],导致生物多样性的严重丧失和生产力下降[6]。
人类活动改变着原有草地生态系统的分布、结构和功能[7,8],草地利用和管理方式的转变常直接影响地表的植被覆盖状况和群落的组成[9]。草地利用管理方式变化造成的地表植被的改变往往具体表现为群落物种结构组成、生物多样性水平和生产力水平变化等群落改变[10]。已往以土地利用变化为驱动和变量的已有研究主要关注于土壤[11,12]、生态系统碳源汇功能[13],这些方面已有较为深入的探讨和报道,但对利用、管理方式改变对群落结构和生产力构成的报道相对较少。同时从植被生态角度对具体人类活动对植物群落的物种组成、多样性和生产力的影响和改变方面的研究,往往各自关注单一驱动因素或单一驱动梯度所造成的群落覆盖改变,如降水[14]、火烧[15-17]、放牧[6,18]、围封[19]、刈割[20,21]和开垦[13]。这些针对性较强的研究虽然对各不同驱动因素的作用机理和生态后果有深入的解析、认识,但却不能充分厘清和比较相近区域内同一自然群落本底在不同人类利用、管理活动作用下生产力和物种组成、多样性发生的变化和差异。
为更为系统和深入的了解和比较不同土地利用、管理方式下,人类活动对典型温性草地群落物种组成、多样性和生产力水平及其构成比例的改变和影响,本研究选取锡林郭勒盟白音锡勒牧场为研究区,采用野外调查、取样测定等群落学调查方法,对6种不同利用、管理方式草地群落类型进行观测描述和比较分析,以期为人类活动对草原自然群落的影响和改变有更深入的理解,为了解草地利用管理方式差异造成的覆被变化所产生的生态意义提供参考。
1 材料与方法
1.1 研究区域概况
本研究于2011年8月(全年群落生物量最大)在中国科学院锡林郭勒草原生态系统国家野外科学观测研究站进行。研究区域范围 N 43°26′~44°08′,E 116°04′~117°05′,海拔1 200~1 300 m,地处白音锡勒草原,内蒙古高原中部,属中纬度西风带半干旱,干旱季风性气候。年均气温-0.4℃,1月和7月份平均气温分别为-23和17.9℃,年均降水量350 mm,主要集中在6-8月,全年大风干燥天气为主[22]。土壤类型以栗钙土为主[23]。
当地地带性气候顶级植被为典型温性草原,如大针茅群落(Form.Stipa grandis)、克氏针茅群落(Form.Stipasareptanavar.krylovii)等。自然群落主要建群植物为大针茅、克氏针茅、羊草(Leymuschinensis)、糙隐子草(Cleistogenessquarrosa)、冷蒿(Artemisiafrigida)等,常见优势种还包括小叶锦鸡儿(Caraganamicrophylla)、冰草(Agropyroncristatum)、米氏冰草(Agropyronmichnoi)、羽茅(Achnatherumsibiricum)、芨芨草(Achnatherumsplendens)等。
1.2 研究方法
1.2.1 样地选取 研究区域内主要植被类型为天然大针茅草地,局地存在部分天然羊草草地。草地主要利用方式为私人放牧场,少量区域为公共放牧场,另有部分草原开垦为人工种植的粟(Setariaitalicavar.germanica)-黍(Panicummiliaceum)人工饲草。研究站于1979年开始对部分大针茅群落和羊草群落开展围封禁牧的研究,进行了围栏封育处理。此外,2005年,大针茅群落部分区域经历草原火灾,该过火区域植被进行了围封恢复处理。
研究采用典型样地取样法,在位置相互接近,非生物环境因子尽可能相似的原则下,除选取区域自然顶级,围封大针茅群落、围封羊草群落作为对照外,依据利用、管理方式的不同分别选择大针茅群落公共放牧区域、大针茅群落私人放牧区域、大针茅群落火烧围封恢复区域以及人工饲草区域,划取建立具有群落代表性的典型样地,以上共计6种样地类型分别记为:1)围封大针茅(FS):围栏封育大针茅群落,建群种大针茅,生物量大,群落组成相对单一;2)围封羊草(FL):围栏封育羊草群落,围封初期羊草为群落主要组成物种,但围封后羊草在群落组成中比例逐渐下降,建群种已转变成羽茅;3)公牧大针茅[24](Pub):大针茅草地公共放牧区域,物种组成较单一,目测植被高度、盖度与地上部分生物蓄积量均显著低于毗邻区域;4)私牧大针茅[24](Pri):大针茅草地私有放牧区域,相对大针茅草地的公牧区域植被状况稍好,仍然显著差于多年围封的大针茅草地。5)火烧围封(FF):经历草原大火后已经围封恢复6年的草地区域,群落内物种丰富,但群落分布并不均匀,群落整体外观呈现斑块状;6)人工饲草(A):由天然草地翻耕开垦,主要种植粟黍人工饲草,每年春季播种,夏末收割作为牲畜饲料使用。
1.2.3 地上及地下生物量测定 利用收获法测定样方地上生物量,在每个样地的5个1 m×1 m样方重复里齐地面刈割样方内植物,并分种放入信封,在烘箱中恒温70℃烘干48 h至恒重后称重记录[25]。地下生物量的测定采用土钻法(直径70 mm)[26-28],在地上部分刈割完毕的每个样方内,按照对角线取土钻3钻,每钻分3层各取10 cm土柱(0~10,10~20,20~30 cm),同样方同层的3份钻样混合作为一个地下生物量样品放入自封袋,即每样地类型分3层取样,每层地下取样品5个。样品放入0.3 mm的网袋,水洗分离土壤和根系,随后用0.3 mm网筛分离石砾等非根杂质,在70℃的烘箱内烘干48 h至恒重并称重。
1.2.4 物种多样性测定与计算方法
1)物种丰富度

2)多样性指数

3)均匀度指数
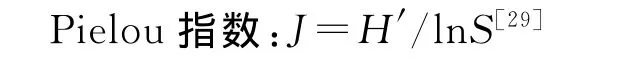
式中,S为物种种数;A为样方面积(m2);Pi为种i的重要值,本研究重要值的计算公式[30]为:

式中,IV为重要值。
通过验收测试的机器人丁达向喵星飞鼠大使提出了释放幽之谷居民的请求,而喵星飞鼠大使则用影态狙击枪扫射居民,以警告丁达不要有非分之想。镜心羽衣怒不可遏,命令丁达一脚踹掉喵星飞鼠大使的枪械。飞鼠群见状,开始疯狂报复。壶天晓为了给丁达争取恢复战斗力的时间,与飞鼠展开了激烈的正面对决。喵星飞鼠大使用大口径超级狙击枪,掳来壶天晓和镜心羽衣的队友当人质。镜心羽衣陷入深深的自责中。
1.3 数据分析
用Excel对数据进行初步的整理和作图,用SPSS 12.0进行数据统计分析。
2 结果与分析
2.1 群落结构特征与物种组成
围封大针茅、私牧大针茅和公牧大针茅具有相近的群落结构和物种组成(表1),建群种大针茅重要值均在0.42~0.44,仅在组成物种上略有差别,随着利用强度的增大,一些数量较少、不耐践踏和采食的物种逐渐消失。人工饲草群落的建群种为粟(IV为0.33),群落内物种组成与其他天然群落迥异,大多为天然草原不会出现的农田杂草。围封羊草群落于围封开始时建群种为羊草 (IV为0.16),但围封后群落逐渐演替变化,群落目前的建群种为羽茅 (IV为0.25)。火烧围封样地群落呈现明显的斑块状分布,为过火后次生裸地基础上恢复的不同群丛类型的混合,样地群落组成尚在变化和重构中,尚未达到稳定。其主要优势种包括冷蒿,木地肤,大针茅(IV分别为0.12,0.13,0.16),群落内组成物种重要值均彼此接近,没有出现IV>0.2的明显建群物种。各群落平均高度和平均盖度的One-way anova结果如表1所示,围封大针茅、人工饲草、围封羊草三者间差异不显著(P>0.05),均具有较高的群落高度(60 cm左右)和较大的群落盖度(近100%)。私牧大针茅在平均高度和盖度上均显著低于围封大针茅(P<0.05)。公牧大针茅群落较之私牧大针茅又存在显著下降(P<0.05)。此外,火烧围封样地的平均高度和平均盖度与其他类型均差异显著(P<0.05)。
2.2 群落生物量的差异
2.2.1 群落地上生物量的构成和差异 大针茅群落围封处理(FS)总地上生物量显著高于2个放牧处理(Pub、Pri)(P<0.05)(图1)。3个围封处理间(FS、FL、FF)无显著差异(P>0.05),但三者中均值最低的围封羊草群落(围封32年),虽显著高于公牧大针茅群落(P<0.05),但与私牧大针茅差异并不显著(P>0.05);火烧围封群落(围封6年),地上平均生物量与2个围封32年处理(FS、FL)间差异不显著,并且以314.48 g/m2的较高均值水平与围封32年大针茅群落相当(313.65 g/m2)。人工饲草植被的地上部分生物量均显著高于其他各利用方式下群落处理,达到586.9 g/m2。
地上生物量的C3和C4植物构成比例在群落之间有着巨大的差异。大针茅群落随着利用强度的降低,C4植物占总地上生物量的比例逐渐减少,其中围封大针茅群落在各群落类型中C4植物比例最低(4%),显著低于两放牧大针茅群落Pub和Pri(P<0.05)(图2)。而人工饲草群落C4植物的比例在各处理中最高,达89%,显著高于各自然群落(P<0.05)。此外,火烧围封群落中C4植物比例亦较高,显著高于其他各自然草地利用类型(P<0.05)。围封大针茅和围封羊草间无显著差异(P>0.05)。
人工饲草群落C4植物地上生物量最大(527.00 g/m2),并显著高于各天然群落类型(P<0.05)。在天然草地中,火烧围封群落的C4生物量增加明显(P<0.05),剩余4种群落类型相互间差异不大(P>0.05)(图2)。与C4植物地上生物量的变化规律不同,随着利用程度的降低,大针茅群落C3植物生物量呈显著增加趋势(P<0.05),围封大针茅C3植物地上生物量在各群落类型最高,达301.76 g/m2,围封羊草群落显著低于围封大针茅群落(P<0.05),而与火烧围封样地差异不大(P>0.05)。

表1 不同利用方式下群落特征及物种组成Table 1 Community characteristics and species composition in different grassland manage type
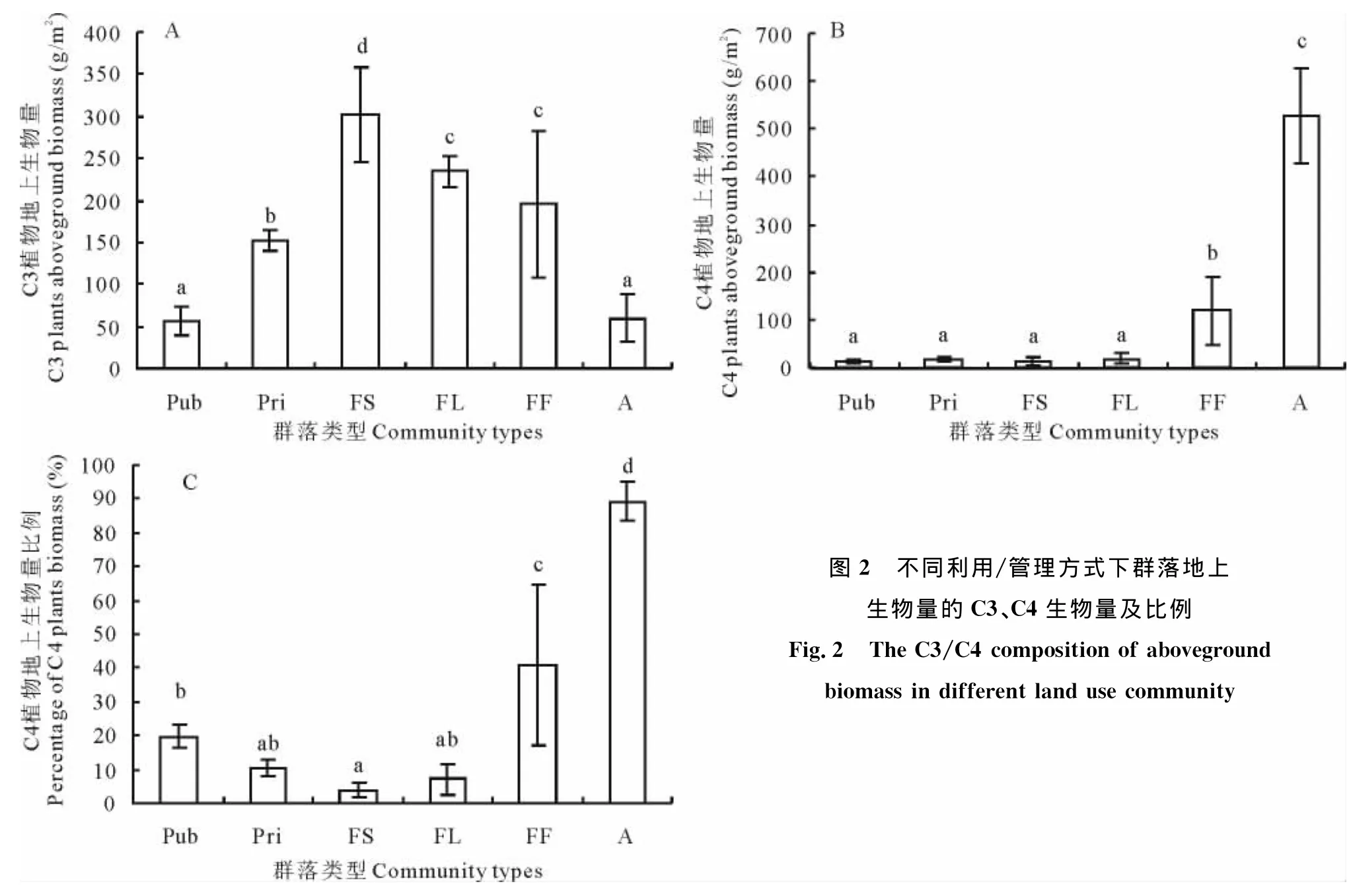
2.2.2 群落地下生物量的组成和差异 大针茅群落随着利用强度的减弱,地下生物量呈上升趋势,在围封大针茅群落达到最大均值(2 943.62 g/m2)(图3)。同是32年围封的大针茅与羊草群落之间并无显著差异(P>0.05),两者均显著高于其他所有群落类型。人工饲草群落总地下生物量在各群落类型中最低(557.00 g/m2),显著低于围封群落FS、FL和FF(P<0.05),但与2个放牧大针茅群落(Pub、Pri)的总地下生物量无显著差异(P>0.05)。
在地下生物量的垂直空间分布上,各群落类型之间差异不大(P>0.05),0~10 cm占总地下生物量(0~30 cm)的50%左右(50.84%±6.24%),10~20 cm约合3层总生物量的30%(30.42%±8.70%),20~30 cm大约只占不到20%(18.73%±3.96%)。人工饲草和放牧大针茅0~10 cm地下生物量显著低于围封大针茅和围封羊草(P<0.05),私牧大针茅和火烧围封群落介于之间,相互间差异不显著(P>0.05)。10~20 cm地下生物量除围封大针茅显著较高外(P<0.05),其余各组间差异不大(P>0.05)。围封羊草群落的20~30 cm地下生物量显著高于放牧大针茅群落(Pub、Pri)和人工饲草群落,火烧围封和围封大针茅群落介于两组之间,与各组差异不显著(P>0.05)。
2.2.3 群落的总生物量和地上地下比 各群落总生物量呈现的规律与地下生物量相近而与地上部分生物量的相对多寡差异较大。围封大针茅与围封羊草彼此接近(P<0.05),显著高于其余4个群落类型(图4)。火烧围封样地显著高于总生物量最低的公牧大针茅样地(P<0.05),私牧场大针茅与人工饲草群落处于两者之间,与两者差异均不显著(P>0.05)。

图1 不同利用(管理)方式下总地上生物量差异Fig.1 Aboveground biomass in different land use(management)community
群落的地上、地下部分比例,在各自然植被间差异不大(P>0.05)(图5),人工群落的地上地下比例特征与自然群落间差异显著(P<0.05),地上部分比例显著高于自然植被,地下部分生物量比例则较之显著降低。在各组自然群落样地内,私牧大针茅群落的地上部分比例稍高于公牧大针茅和围封大针茅,围封羊草和围封大针茅彼此间非常接近,而火烧围封群落的地上部生物量比例也稍高。
2.3 物种多样性
群落的物种丰富度是群落的重要特征[6],在3个放牧与围封的大针茅-糙隐子草群落(Pub、Pri和FS)中,公牧大针茅群落物种丰富度均值稍低于私牧和围封的大针茅群落,但三者间并无显著差异(表2)。3个大针茅群落的物种丰富度均显著低于围封羊草,火烧围封和人工饲草群落。围封羊草群落的物种丰富度最高(11.2±1.924),显著高于其他5个群落类型。人工饲草与火烧围封群落间无显著差异。
物种多样性和均匀度水平一定程度反映了群落的稳定性和发育阶段,一般认为群落多样性和均匀度水平越高群落就越稳定[31,32]。样地水平的Gleason指数和Shannon-Wiener指数大致呈现相同的规律:随着利用程度的降低,大针茅群落在Pub、Pri和FS间逐渐上升,但三者间差异并不显著(P>0.05);围封羊草和火烧围封群落的物种多样性水平显著高于人工饲草群落和各大针茅群落(P<0.05),两者间差异不显著(P>0.05)。而人工饲草群落物种多样性水平显著高于各大针茅群落(P<0.05)(表2)。
围封大针茅和私牧大针茅群落具有显著较低的群落均匀度水平,火烧围封样地的均匀度水平最高(0.929±0.026),显著高于3个不同管理方式不同利用程度的大针茅群落(P<0.05)。公牧大针茅、围封羊草和人工饲草间差异不显著(P>0.05),围封羊草、火烧围封和人工饲草间差异不显著(P>0.05)。这反映了较低水平的利用程度或对大针茅群落的围封保护使群落内物种重要值的差异逐渐变大,优势种的优势程度被增强,群落的均匀程度降低[33]。而火烧围封中的FF群落则不然,火烧产生的空缺生态位和近乎次生裸地的充裕生存空间给竞争能力各不相同的草地物种创造低竞争压力的生存机会,从而群落内各种的重要值和群落地位差异并未充分显露和稳定,使得群落的均匀度呈现较高水平。
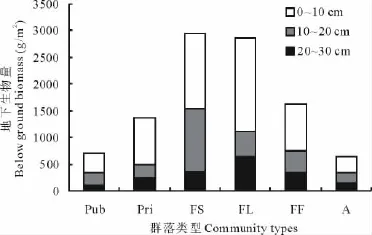
图3 不同利用/管理方式对群落地下生物量及其分层构成的影响Fig.3 The below-ground biomass in different land use community and below-ground biomass in three depth layers
图4 不同利用/管理方式下群落总生物量Fig.4 Total biomass in different land use community
3 讨论
3.1 草地利用管理方式对草地群落特征、物种组成和多样性的影响

图5 不同利用/管理方式下群落地上、地下生物量所占比例Fig.5 The percentage of above/below ground biomass in different land use community
草地的不同利用管理方式中,以开垦和火烧对物种组成、多样性的影响最为显著。开垦形成的人工饲草群落虽然较原有的大针茅群落具有更高的物种多样性水平(表2),但是人工植被和天然植被的组成物种截然不同(表1)。火烧围封处理很大程度丰富了大针茅群落的物种组成,显著提高了群落的物种多样性水平,群落的物种丰富度增至与围封羊草群落大致相当。与此同时,群落的盖度、高度特征也发生很大改变,这些变化与周道玮和刘仲龄[15]对羊草群落的火烧后果研究相似。围封大针茅和围封羊草围封时间相同,环境因子相似,两者间的物种组成和多样性的差异是群落类型差异的结果,大针茅群落相对羊草群落来说,物种组成更加单一[34-36]。3个大针茅群落Pub、Pri和FS之间相似的物种组成反映围封、禁牧与否以及放牧程度的差异对群落物种组成的影响相对较小[19,37],但是围封与较低的放牧水平显然会提高群落的盖度、高度特征,影响了群落的整体外观。
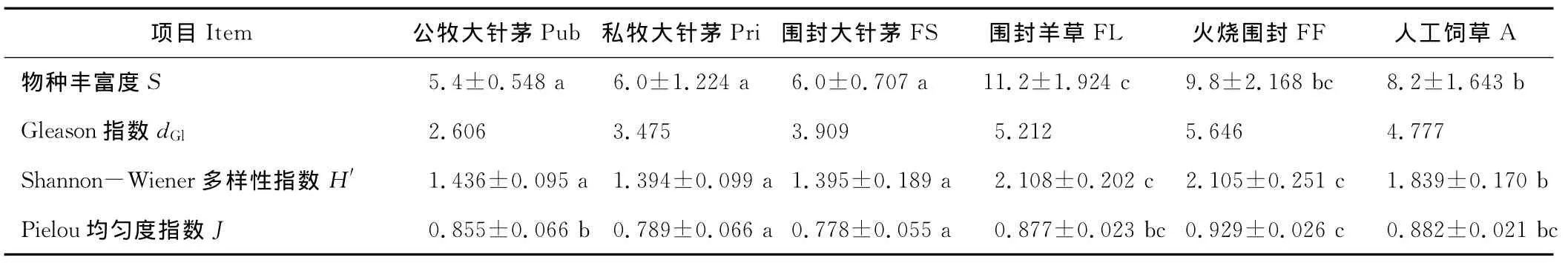
表2 不同利用方式下群落物种多样性特征Table 2 Community species diversity in different grassland manage type
3.2 草地利用(管理)方式对草地生产力的影响
草地利用管理方式差异对群落生产力也产生着重要的影响[13,38]。围封被认为逐年增加群落的地上生物量[39],放牧的大针茅群落(Pub、Pri)与围封大针茅群落间生产力的比较结果支持以上观点。更细致的生产力拆分比较结果进一步说明大针茅群落在不同管理方式下生产力构成上的改变,随着利用程度的降低群落地上生物量显著增加的部分主要由C3植物的产量增长贡献,C4植物的生物量未明显增加,其比例沿着草地利用程度降低逐渐下降。因为C4植物相对C3植物有着更高的水分利用效率[40],这一构成改变可能会潜在影响群落水平的水分利用效率[41],即利用程度越低,WUE可能反而逐渐下降。围封时间相同的大针茅和羊草群落(32年)与围封时间较短的FF群落(6年)地上生产力差异不显著(图1),可见一定围封时间之后,围封时间的长短不再是影响群落地上生产力水平的主要因素。
此外,一定的火烧可能对群落地上生产力水平有着某种促进作用。火烧围封群落C4植物的产量和比例较其他自然群落(除人工饲草群落外)均显著增高,可见火烧对群落物种组成、生产力构成的改变,即过火后C4植物生物量的增多可能是促使群落获得较高生产力的原因。
草原开垦曾被认为在释放土壤碳素[42]的同时,亦会改变群落生物量的地上地下比[42,43]。比较围封大针茅群落,人工饲草群落间差异表明,开垦使得群落地上生物量显著增加,但地下生物量损失严重,虽然显著增加了群落的地上地下比,但是群落总生物量仍明显损失,但最终总生物量与放牧大针茅群落(Pub,Pri)差异并不大(P>0.05,图4)。人工饲草地上部分生物量组成中C4植物无论从比例(图6)还是绝对产量(图2B)上都与天然植被差异巨大。高比例的栽植C4植物作为替代饲草,相较天然植被显著提高了草地的地上生产力,同时预计也将大幅度提高群落的水分利用效率[44]。
4 结论
区域的调查结果表明,围封大针茅群落与围封羊草群落生物量及其组成相近,但物种多样性水平较低。围封会增加大针茅地上、地下生物量,但对物种组成和多样性的影响不大,而放牧造成的退化主要减少了群落的地上地下生物量,以地下生物量的损失更为严重,同时C3植物的生物量损失相对C4更为严重,C3植物在群落地上生物量的构成比例逐渐下降。火烧显著丰富了群落的物种组成,增加了群落物种多样性,但对群落的盖度、高度和生产力水平的损失作用明显。人工饲草地的开垦彻底改变了天然群落的物种组成,物种多样性水平近于天然羊草群落而高于大针茅群落,同时地上生物量显著增加,但由于地下生物量损失严重,群落总生产力相对围封自然群落显著降低,近于公牧的天然大针茅群落。
[1] Scurlock J,Hall D.The global carbon sink:a grassland perspective[J].Global Change Biology,1998,4(2):229-233.
[2] 胡中民,樊江文,钟华平,等.中国草地地下生物量研究进展[J].生态学杂志,2005,24(9):1095-1101.
[3] Fan J,Zhong H,Harris W,etal.Carbon storage in the grasslands of China based on field measurements of above-and belowground biomass[J].Climatic Change,2008,86(3):375-396.
[4] 呼格吉勒图,杨劼,宝音陶格涛,等.不同干扰对典型草原群落物种多样性和生物量的影响[J].草业学报,2009,18(3):6-11.
[5] 李博.中国北方草地退化及其防治对策[J].中国农业科学,1997,30(6):1-9.
[6] 汪诗平,李永宏.不同放牧率对内蒙古冷蒿草原植物多样性的影响[J].植物学报:英文版,2001,43(1):89-96.
[7] 高志强,刘纪远,曹明奎,等.土地利用和气候变化对农牧过渡区生态系统生产力和碳循环的影响[J].中国科学:D辑,2004,34(10):946-957.
[8] Turner B,Skole D,Sanderson S,etal.Land-use and land-cover change.Science/Research Plan[J].Global Change Report,1995,35:108-123.
[9] 苗福泓,郭雅婧,缪鹏飞,等.青藏高原东北边缘地区高寒草甸群落特征对封育的响应[J].草业学报,2012,21(3):11-16.
[10] 李秀彬.全球环境变化研究的核心领域[J].地理学报,1996,51(6):553-558.
[11] 傅伯杰,陈利顶,马克明.黄土丘陵区小流域土地利用变化对生态环境的影响[J].地理学报,1999,54(3):241-246.
[12] Fu B,Gulinck H,Masum M.Loess erosion in relation to land-use changes in the Ganspoel catchment,central Belgium[J].Land Degradation &Development,1994,5(4):261-270.
[13] 李凌浩.土地利用变化对草原生态系统土壤碳贮量的影响[J].植物生态学报,1998,22(4):300-302.
[14] 武建双,李晓佳,沈振西,等.藏北高寒草地样带物种多样性沿降水梯度的分布格局[J].草业学报,2012,21(3):17-25.
[15] 周道玮,刘仲龄.火烧对羊草草原植物群落组成的影响[J].应用生态学报,1994,5(4):371-377.
[16] Daubenmire R.Ecology of fire in grasslands[J].Advances in Ecological Research,1968,5:209-266.
[17] Whelan R J.The Ecology of Fire[M].Cambridge:Cambridge University Press,1995:15-18.
[18] 王艳芬,汪诗平.不同放牧率对内蒙古典型草原地下生物量的影响[J].草地学报,1999,7(3):198-203.
[19] 左万庆,王玉辉,王风玉,等.围栏封育措施对退化羊草草原植物群落特征影响研究[J].草业学报,2009,18(3):12-19.
[20] 仁青吉,罗燕江,王海洋,等.青藏高原典型高寒草甸退化草地的恢复——施肥刈割对草地质量的影响[J].草业学报,2004,13(2):43-49.
[21] 马涛,武高林,何彦龙,等.青藏高原东部高寒草甸群落生物量和补偿能力对施肥与刈割的响应[J].生态学报,2007,(6):2288-2293.
[22] 赵雪艳,汪诗平.不同放牧率对内蒙古典型草原植物叶片解剖结构的影响[J].生态学报,2009,29(6):2906-2918.
[23] 王艳芬,陈佐忠,Tieszen L T.人类活动对锡林郭勒地区主要草原土壤有机碳分布的影响[J].植物生态学报,1998,22(6):545-551.
[24] 苏大学,张自和,陈佐忠,等.天然草地退化、沙化、盐渍化的分级指标[Z].GB 19377-2003,2004.
[25] 樊江文,杜占池,钟华平.红三叶,鸭茅生物量和叶面积时空结构特征[J].草地学报,2004,12(3):204-208.
[26] 宇万太,于永强.植物地下生物量研究进展[J].应用生态学报,2001,(6):927-932.
[27] 宋璐璐,樊江文,钟华平,等.重庆红池坝地区草地群落生物量及物种丰富度沿海拔梯度格局变化[J].草地学报,2010,18(2):160-166.
[28] 刘凤山,周智彬,胡顺军,等.根钻不同取样法对估算根系分布特征的影响[J].草业学报,2012,21(2):294-299.
[29] Magurran A E.Ecological Diversity and its Measurement[M].Princeton,NJ:Princeton University Press,1988:179.
[30] 孙菊,李秀珍,胡远满,等.大兴安岭沟谷冻土湿地植物群落分类,物种多样性和物种分布梯度[J].应用生态学报,2009,20(9):2049-2056.
[31] Tilman D,Downing J A.Biodiversity and stability in grasslands[J].Ecosystem Management:Selected Readings,1996,367:363-365.
[32] Baskin Y.Ecosystem function of biodiversity[J].BioScience,1994,44(10):657-660.
[33] 康永祥,康博文,岳军伟,等.陕北黄土高原辽东栎 (Quercusliaotungensis)群落类型划分及其生态位特征[J].生态学报,2007,10:4096-4105.
[34] 王玉辉,何兴元.放牧强度对羊草草原的影响[J].草地学报,2002,10(1):45-49.
[35] 白永飞,陈佐忠.锡林河流域羊草草原植物种群和功能群的长期变异性及其对群落稳定性的影响[J].植物生态学报,2000,24(6):641-647.
[36] 李永宏.放牧影响下羊草草原和大针茅草原植物多样性的变化[J].植物学报,1993,35(11):877-884.
[37] Meissner R A,Facelli J M.Effects of sheep exclusion on the soil seed bank and annual vegetation in chenopod shrublands of South Australia[J].Journal of Arid Environments,1999,42(2):117-128.
[38] 白永飞,李凌浩,王其兵.锡林河流域草原群落植物多样性和初级生产力沿水热梯度变化的样带研究[J].植物生态学报,2000,24(6):667-673.
[39] 单贵莲,徐柱,宁发,等.围封年限对典型草原群落结构及物种多样性的影响[J].草业学报,2008,17(6):1-8.
[40] Tanner C B,Sinclair T R.Efficient water use in crop production:research or re-search[A].Limitations to Efficient Water Use in Crop Production[M].1983:1-27.
[41] 胡中民,于贵瑞,王秋凤,等.生态系统水分利用效率研究进展[J].生态学报,2009,29(3):1498-1508.
[42] Anderson D,Coleman D C.The dynamics of organic matter in grassland soils[J].Journal of Soil and Water Conservation,1985,40(2):211-216.
[43] McConnell S,Quinn M L.Soil productivity of four land use systems in southeastern Montana[J].Soil Science Society of A-merica Journal,1988,52(2):500-506.
[44] Sinclair T R,Tanner C,Bennett J.Water-use efficiency in crop production[J].BioScience,1984,34(1):36-40.
