60Co~γ射线对库尔勒香梨的辐射效应
2013-04-04覃伟铭何诚诚
尹 航 ,覃伟铭 ,南 鑫 ,杨 振 ,何诚诚 ,李 疆
(1.新疆农业大学 林学与园艺学院,新疆 乌鲁木齐 830052;2.新疆巴音郭楞蒙古自治州沙依东园艺场,新疆库尔勒 841000;3. 新疆巴音郭楞蒙古自治州库尔勒市园林绿化管理局,新疆 库尔勒 841000;4. 新疆伊犁自治州察布查尔县林业局,新疆 伊犁 835000)
库尔勒香梨皮薄肉脆,清甜爽口,细嫩多汁,以其独特的芳香最为出名。库尔勒香梨属蔷薇科梨属植物,在新疆的栽培历史悠久,在国际市场上享有盛誉,深受国内外消费者的喜爱[1-3]。近年来,库尔勒香梨存在的一些问题影响了经济效益的继续扩大,例如果实可食率较低、果形不一致、宿萼等,因此亟需进一步对库尔勒香梨果实品质进行改善,并研究培育库尔勒香梨新品种[4-5]。
在生产中,人们对品种的改良以及新品种发现与创造,用到的方法包括引种、实生选种、芽变选种、杂交育种、辐射诱变育种以及利用生物技术等[6-8]。在梨的研究中,生物技术应用增多,尤其在种质资源基因鉴定方面[9-11]。但是,辐射诱变育种可以方便快捷并大量得到新品种,所以在对植物变异的研究中仍得到广泛应用[8]。辐射诱变存在变异谱广、突变率高等特点,在创造新种质、选育新品种研究中是一种有效途径[12]。辐射诱变育种多为利用各种射线对植物的枝条、种子、花粉进行照射,如X射线、γ射线、β射线、中子、激光、电子束、离子束、紫外线等,从而可获得大量新的种质资源以及新的品系[13-15]。60Co~γ射线是较常用的辐射源,可对植物的生长发育产生一定的抑制或促进作用,在香蕉[16-17]、沙田柚[18-19]、梨[20-22]、苹果[23]、草莓[24]、黑莓[25]等水果的研究中均有应用。本试验中从2011年开始,采用60Co~γ射线辐照诱变技术,对库尔勒香梨1年生休眠枝和萌动枝进行诱变处理[21],观察2011年和2012年60Co~γ射线对库尔勒香梨枝条的影响,并在2012年对成活的辐射枝跟踪观察,旨在改善库尔勒香梨果实品质,进一步研究培育库尔勒香梨新品种。
1 材料与方法
1.1 材 料
试验中用于辐射处理的库尔勒香梨枝条均采自于新疆巴州库尔勒市沙依东园艺场,2011年3月20日前剪取休眠枝,2011年4月4日前剪取萌动枝。各梯度辐射处理约30根枝条,长度约为15 cm,每根枝条约带3个饱满芽。辐射处理后,将处理后的枝条再次嫁接到园艺场12年生库尔勒香梨树上。
1.2 方 法
1.2.1 辐射处理
2011年4月6日,在新疆农业科学院辐照中心对所采库尔勒香梨枝条进行60Co~γ射线辐射处理,以未进行辐射处理的枝条为对照组(CK)。对休眠枝的处理分别为0 Gy(X0)、20 Gy(X20)、30 Gy(X30)、40 Gy(X40)、50 Gy(X50)、60 Gy(X60)共6组;萌动枝的处理分别为0 Gy(M0)、20 Gy(M20)、30 Gy(M30)、40 Gy(M40)共 4组,剂量率为1 Gy/min。
1.2.2 观测方法
2011年4月,辐射枝嫁接后,开始对其变异情况进行观察,主要观察叶片和枝条的外部形态变异情况以及物候期的变化情况。
2012年4月,辐射枝嫁接第2年,对辐射枝的物候期和花朵雌蕊、雄蕊数量进行观测,并对第2年新发枝长度和粗度、叶片长度和宽度、叶片面积(便携式手持叶面积仪CI-203)和变异情况进行观测。
2012年4月,辐射枝有开花坐果现象,在对果实各特征指标(果实横纵径、果柄长粗)测量后,于当年9月在试验室内对成熟的果实进行品质测定,主要包括果心大小(游标卡尺测量)、单果鲜质量(电子天平)、果点密度(目测计数)、果实硬度(GY-3型果实硬度计)、可溶性固形物(阿贝折光仪)、抗坏血酸(2,6-二氯酚靛酚滴定法)、可滴定酸(氢氧化钠中和滴定法)、可溶性糖(蒽酮试剂比色法)。因第1年的未辐射枝在2012年时未坐果,在果实坐果率和果实品质测定中,选用与辐射枝同株的库尔勒香梨果实做对照。
1.2.3 数据分析
采用Excel 2007对试验数据进行初步汇总处理,利用SPSS17.0软件对数据进行方差分析,分析方法采用邓肯氏新复极差法(SSR)。
2 结果与分析
2.1 辐射对库尔勒香梨枝条嫁接成活率的影响
辐射对库尔勒香梨枝条嫁接成活率的影响见表1,2012年休眠枝的存活量要多于萌动枝的存活量。从表1中可以看到,第1年萌动枝的嫁接成活率随着辐射剂量的升高而增大,表明低剂量的辐射刺激有助于提高萌动枝的嫁接成活率。第1年的休眠枝的嫁接成活率随着辐射剂量的升高有降低的趋势,仅有X20处理组的成活率比对照组高。表明低剂量辐射对提高休眠枝条嫁接成活率有一定的刺激作用,但总体随着辐射剂量的提高,对枝条造成的损伤加重,抑制枝条的成活。
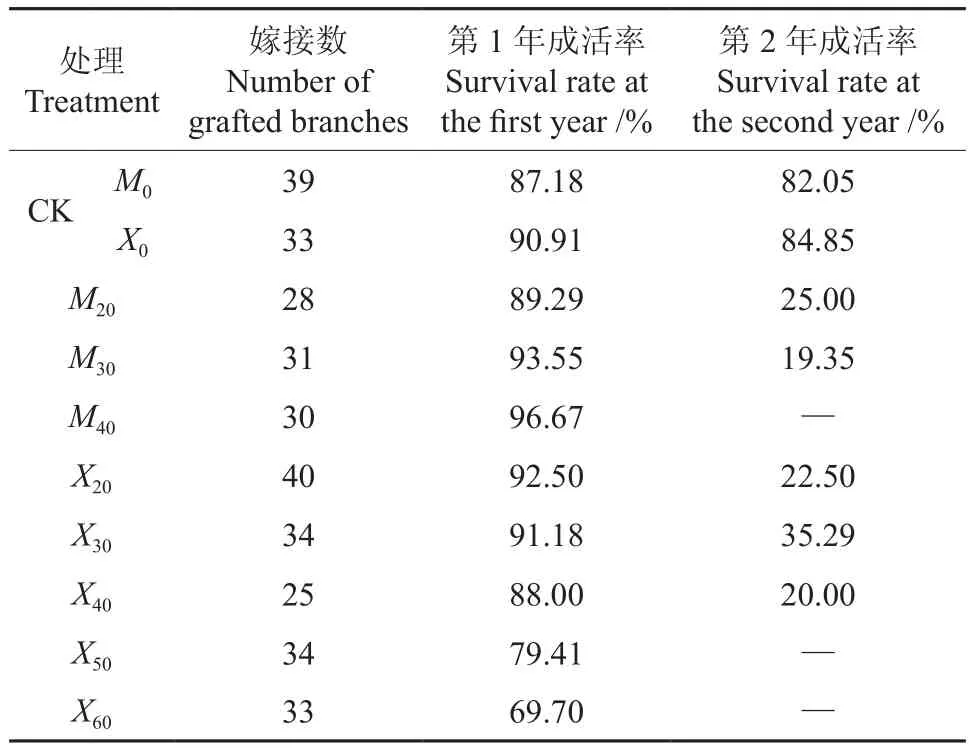
表1 辐射对库尔勒香梨枝条嫁接成活率的影响Table 1 Effect of radiation on survival rate of grafted branches in Korla Fragrant Pear
在第2年对辐射枝的观测中发现,总体枝条嫁接成活率有随着辐射剂量的提高而下降的趋势。其中X30处理组的成活率要高于其它处理。M40、X50、X603组在第2年未能发出新梢,只留有第1年的短粗枝。通过表1可以看出,小剂量辐射有利于提高嫁接枝条的成活率,30 Gy剂量的辐射对休眠枝的成活率影响较大。
2.2 辐射对库尔勒香梨物候期的影响
辐射对库尔勒香梨物候期的影响见表2。在观察中发现,处理M0和X0的物候期相同,故将二者合并作为对照与辐射枝进行对比。从表2中可以看出,2011年辐射枝仅有展叶、抽枝,未能开花结果,辐射枝的物候期相对于对照有延迟现象,并随辐射剂量的增大,物候期延迟增长。2012年,辐射枝出现开花、坐果现象,对照组虽有开花,但未坐果。2012年,在对辐射枝物候期的观察中可以看出,辐射枝的花期晚于对照,并随着辐射剂量的增高,花期有延迟现象,但是辐射枝与对照的展叶期、新梢生长期、坐果期和果实成熟期大体相同。通过对物候期的观察,发现辐射枝在第2年出现变异回归现象,辐射效应开始减弱。

表2 辐射对库尔勒香梨物候期的影响(日/月)Table 2 Effect of radiation on phenophase of Korla Fragrant Pear (D/M)
2.3 辐射对库尔勒香梨不同器官的影响
2.3.1 辐射对库尔勒香梨叶片的影响
第1年变异情况:异形叶所占比例较大,随着剂量的升高变异率升高,存在大量的变异叶。20 Gy时变异率达到30%,高剂量辐射枝所发新叶大部分为异形叶,60 Gy时变异率达到100%,叶片形态有半叶、两尖叶、叶片缺刻、波状叶缘等变异(见图1)。

图1 2011年叶片形态变异情况Fig.1 Morphological variation status of leaves in 2011
第2年变异情况:仅有几片叶发生较大的形态变异,其余叶片部分有叶型及叶缘锯齿的变化,包括叶平基、锯齿变小。田间观察发现,大部分叶片与对照组正常叶片在外部形态上无明显差异(见图2)。

图2 2012年叶片形态变异情况Fig.2 Morphological variation status of leaves in 2012
2012年,对成熟叶片进行测量,测定结果如表3所示。由表3可知,随着辐射剂量的升高,休眠枝叶片长度和宽度变大,且X40、X30组的叶片长度显著大于X20组;X40组的叶片宽度极显著大于X30、X20组。而萌动枝中,M30组的叶片长度显著小于M20组,但叶片宽度差异不显著。M30组的叶片长度极显著小于CK。结果表明,辐射后的第2年,休眠枝的叶片长度增大,萌动枝的叶片长度减小,小剂量辐射对辐射枝的叶片宽度影响不显著。
CK的叶面积极显著大于辐射枝,辐射枝叶面积在40~50 cm2之间。休眠枝的叶面积随着辐射剂量的升高而增大,萌动枝的叶面积随着辐射剂量的升高而降低。X40组的叶面积极显著大于X30、X20组,X30和X20组叶面积差异不显著。M20组叶面积显著大于M30组。结果表明,辐射后的第2年,辐射枝的叶片面积有减小现象。

表3 2012年辐射对库尔勒香梨叶片性状的影响†Table 3 Effect of radiation on leaf character in Korla Fragrant Pear in 2012
2.3.2 辐射对库尔勒香梨枝条的影响
第1年变异情况:新梢的变异率随着辐射剂量的提高而增大,高剂量辐射枝新梢变异率极高,20 Gy时变异率为35%,60 Gy时可达到100%,形态出现分叉枝、丛状矮枝等。枝条形态变异情况见图3。

图3 2011年枝条形态变异情况Fig.3 Morphological variation status of branches in 2011
第2年变异情况:新梢形态多丛状枝、分叉枝等形状变异,有少部分粗短枝。多数枝条细长,长势较弱,少部分枝条长势旺盛,粗长。田间观察结果表明,在第2年,新梢生长较细弱者,多为挂果枝条,长势旺盛的辐射枝极少挂果,甚至未挂果。枝条形态变异情况见图4。

图4 2012年枝条形态变异情况Fig.4 Morphological variation status of branches in 2012
2012年辐射对库尔勒香梨枝条性状的影响见表4。从表4可以看出,在2012年辐射枝的新梢生长中,随着辐射剂量的升高,辐射枝的新梢长度增大,其中休眠枝长度要大于对照,萌动枝长度小于对照。其中X40和X30组新梢长度显著大于对照,M20组新梢长度显著小于对照。休眠枝的新梢长度显著大于萌动枝的新梢长度。结果表明,辐射枝嫁接后第2年,辐射的刺激对休眠枝新梢长度的生长有促进作用,对萌动枝新梢长度的生长有抑制作用。
在2012年对嫁接枝条新梢粗度的调查中,发现萌动枝的新梢粗度小于对照,且M30组的新梢粗度显著小于对照。休眠枝新梢粗度大于对照,X40和X30组的新梢粗度显著大于对照。休眠枝新梢粗度显著大于萌动枝新梢粗度。结果表明,嫁接后第2年,辐射的刺激促进了休眠枝新梢粗度的生长,抑制了萌动枝新梢粗度的生长。

表4 2012年辐射对库尔勒香梨枝条性状的影响Table 4 Effect of radiation on branch characters of Korla Fragrant Pear in 2012
从表4中可看出,辐射枝的枝间距大于对照,其中X40、X30、M20这3组处理的枝间距显著大于对照。结果表明,受到辐射的影响,枝条的枝间距增大。
2.3.3 辐射对库尔勒香梨花的影响
第1年辐射枝未开花。第2年辐射枝开花结果。
第2年,各处理中库尔勒香梨花的雌雄蕊数量如图5所示。由图5可见,雌蕊柱头数量无差异,均为5个,与正常库尔勒香梨花雌蕊柱头数相同。

图5 辐射对库尔勒香梨雌蕊、雄蕊数量的影响Fig.5 Effect of radiation on pistil and stamen number in Korla Fragrant Pear
辐射枝花丝数量极显著大于对照。萌动枝组内花丝数量无显著差异。休眠枝花丝数量有随着辐射剂量提高而增多的趋势。X40组花丝数量平均达到26.25,显著多于X20组。
2.3.4 辐射对库尔勒香梨果实的影响
(1)辐射对库尔勒香梨果实坐果率的影响
辐射对库尔勒香梨果实坐果率的影响如表5所示。由表5可知,2012年辐射枝嫁接后第2年开始坐果,由于受第2年辐射枝的成活率的影响,M20组开花总数较少,结果量也少。M30组早期坐果率虽然达到45%,但是晚期坐果率较小,结果量少。X40组开花总数较多,但早期坐果率较低,并且在生长过程中,随着自然落果,在成熟期采摘时X40组的果实已经全部脱落。X20组和X30组的早期花朵坐果率和晚期花朵坐果率均较高。
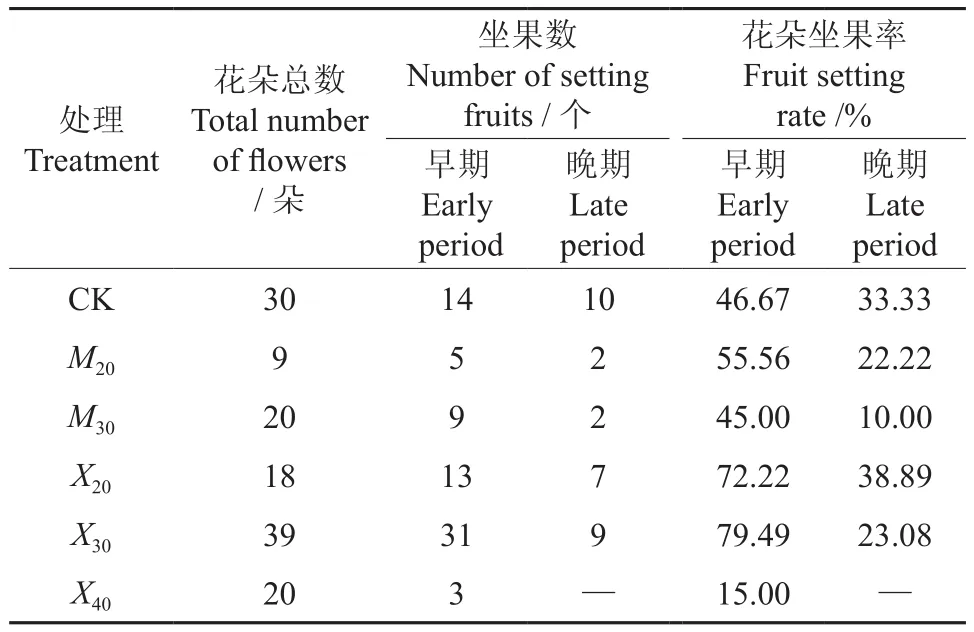
表5 辐射对库尔勒香梨果实坐果率的影响Table 5 Effect of radiation on fruit setting rate in Korla Fragrant Pear
(2)辐射对库尔勒香梨果实外在品质的影响
辐射对库尔勒香梨果实外在品质的影响如表6所示。由表6可知,枝条受辐射后,果实横径、果实纵径、果柄粗度均有随剂量升高而提高的趋势。萌动枝上所结果实的横径、纵径和单果质量均显著大于对照,但果柄粗度、长度与对照差异不显著。X30组在果实横径、果柄粗度上显著大于对照。X20组的各项数据与对照差异均不显著。结果表明,辐射较易促进果实横径生长,30 Gy剂量的辐射会促使果柄变粗。辐射枝上所结果实的性状表现与对照差异不显著。在本试验中,辐射对果实外在品质的综合影响效应不显著,仅在一定程度上增大了果实横纵径和单果质量。
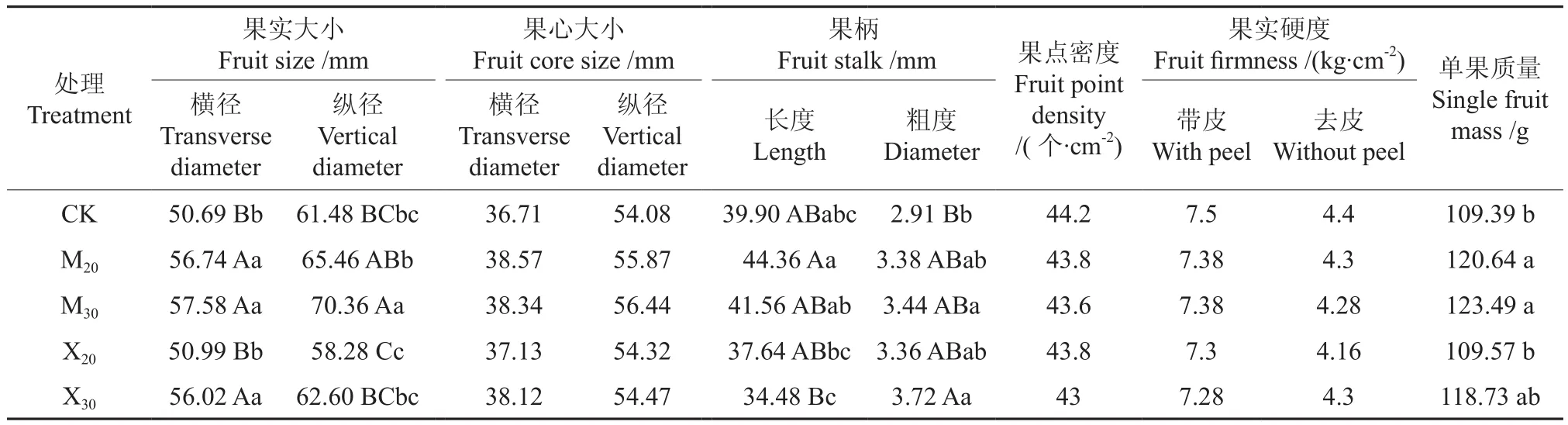
表6 辐射对库尔勒香梨果实外在品质的影响Table 6 Effect of radiation on external fruit quality in Korla Fragrant Pear
(3)辐射对库尔勒香梨果实内在品质的影响
辐射对库尔勒香梨果实内在品质的影响如表7所示。由表7可知,辐射可促进果实中糖含量的提高,且与辐射剂量成正比例关系。X30组的可溶性固形物和可溶性糖含量显著大于对照。萌动枝果实的可溶性糖含量亦显著大于对照。 抗坏血酸含量有减少的趋势,M30组的抗坏血酸含量显著低于对照。从表7中看出,辐射对于果实的含水量和可食率无明显的影响。辐射后果实中滴定酸含量有减少趋势,但差异不显著。因此,辐射枝果实的糖酸含量比有较大改善,X30和M30组的糖酸比显著大于对照。结果表明,辐射有助于果实糖含量的提高,但有减少果实中抗坏血酸含量的趋势。

表7 辐射对库尔勒香梨果实内在品质的影响Table 7 Effect of radiation on interior fruit quality of Korla Fragrant Pear
3 结论与讨论
2011年,枝条受到辐射后,小剂量辐射对枝条的嫁接成活率有一定的提高,但从总体上看辐射会损伤枝条,降低枝条的嫁接成活率[17]。2012年,辐射枝的成活率减少,50、60 Gy剂量辐射的休眠枝和40 Gy剂量辐射的萌动枝均未能发出新芽。且辐射促进了休眠枝新梢长度、粗度的增长,抑制了萌动枝长度、粗度的生长。受辐射影响,辐射枝的枝间距增大,叶片面积减小,符合60Co~γ射线对植物生长的抑制和促进效应[13-14],亦与刘军丽等在观赏海棠以及张锐等对香梨的诱变效应上的研究结果相似[23,26]。但田间观察结果表明,辐射枝的新梢和叶片的外部形态与正常库尔勒香梨相差不大。
2011年,枝条生长受到辐射的影响较大,表现在新梢、叶片的高变异率,以及物候期的延迟[21]。在2012年辐射枝的物候期除在花期上有一定延迟效应外,展叶期、新梢生长期、坐果期及果实成熟期与正常库尔勒香梨物候期无明显差异。
综合枝条、叶片的形态和物候期观察,2012年辐射对枝条生长的影响开始减弱,开始出现变异回归现象。
通过对2012年辐射枝果实外在品质的测定,发现辐射对果实外在品质的综合影响效应较小,在一定程度上增大了果实横纵径和单果质量。在对果实内在品质的测定中,发现辐射提高了果实的可溶性固形物含量、可溶性糖含量和糖酸比,未能提高果实抗坏血酸含量,对可滴定酸的含量影响不大。果实大小和含糖量与乐文全等人的研究结果相似[27]。 综合果实品质的测定结果表明,辐射对果实的影响既有有利因素,亦有不利因素。
综上所述,试验中第1年辐射对库尔勒香梨有较强的变异影响。在枝条嫁接后第2年,辐射效应开始减弱。辐射枝嫁接后第2年便出现坐果现象,果实品质可能尚未稳定,但通过综合表现可以看出,30 Gy剂量的辐射对库尔勒香梨的综合影响较强,有利于库尔勒香梨新品种的选育。
参考文献:
[1] 刘 玮. 库尔勒香梨研究进展[J]. 安徽农学通报,2009,15(11):163-164.
[2] 高启明,侯江涛,李 疆,等. 库尔勒香梨生产现状与研究进展[J]. 中国农学通报, 2005, 21(2): 233-236.
[3] 高启明,李 疆,李 阳,等. 库尔勒香梨研究进展[J]. 经济林研究 , 2005, 23(1): 79- 82.
[4] 王杰君. 库尔勒香梨新品种(系)简介[J]. 西北园艺, 2001,(1): 36 - 37.
[5] 张 燕,吴国良. 目前我国果业发展存在的问题、对策及趋势[J]. 山西果树 , 2003,(3):32 - 33.
[6] Li Xiugen, Yang Jian, Wang Long,et al.Progress and proposal in research of pear breeding in China [J]. Journal of Fruit Science,2010, 27(6): 987 - 994.
[7] 徐刚标. 中国林木遗传改良研究现状[J]. 经济林研究,2003,21(4): 120 - 122.
[8] 徐冠仁. 植物诱变育种学[M]. 北京:中国
[9] 乌云塔娜,包梅荣,李铁柱,等. 中国梨新SFBB-gamma基因的鉴定及序列分析[J]. 中南林业科技大学学报, 2010, 30(8):5-10, 40.
[10] 邓建军,谭晓风,包梅荣,等. 11个中国梨品种S基因型的鉴定[J]. 中南林业科技大学学报,2010,30(3): 63-66.
[11] 许靖诗,乌云塔娜,叶生晶,等. 梨种质资源SSR引物的筛选与评价[J]. 中南林业科技大学学报,2012,32(7): 80-85.
[12] 万国贤. 辐射诱变在创造种质资源上的应用[J]. 作物研究,1994, 8(2): 1 - 4.
[13] Chen Qiufang, Wang Min, He Meimei,et al. Advances inInduced Mutation Breeding by Radiation for Fruit Plants [J].Chinese Agricultural Science Bulletin, 2007, 23(1): 240 - 243.
[14] 王旭军,吴际友,程 勇,等. 辐射育种及其在林木育种中的应用前景[J]. 湖南林业科技, 2007, 34(2): 13-15,18.
[15] 龚洪恩,吕芳德. 离子束生物技术在植物育种中的应用[J].经济林研究, 2008,26(1): 113-116, 121.
[16] Xu Chunxiang, Xie Lijun, Chen Houbin,et al. Effect of60Coγ-Rays irradiation on somatic embryogenesis ofMusaAAA CAVENDISH cv. ‘Baxijiao’ [J]. Journal of Nuclear Agricultural Sciences, 2011, 25(1): 0048 - 0052.
[17] 叶春海,丰 锋,吕庆芳,等. 香蕉60Co辐射诱变效应的研究[J]. 西南农业大学学报:自然科学版,2000,22(4): 301-303.
[18] 黄建昌,肖 艳,赵春香,等. 少核沙田柚的辐射选育研究[J]. 核农学报 , 2003, 17(3): 171 - 174.
[19] 李 庄,陈茂鑫,韦文楼. 无核沙田柚的RAPD研究-无核机理研究(Ⅱ)[J]. 激光生物学报, 2000, 9(1): 4-7.
[20] Liu Chao, Wang Xiaoming, Wang hong. Inhibition effect of radiation on ppo’s activity in pera juice [J]. Journal of Nuclear Agricultural Sciences, 2006, 20(4): 331 - 332.
[21] 杨 振,李 疆,梅 闯,等.60Co-γ辐照对库尔勒香梨枝条当代诱变效应初报[J]. 新疆农业科学,2012,49(5): 848-855.
[22] 王妍炜,叶振风,衡 伟,等. 砀山酥梨60Coγ射线辐射诱变效应及其RAPD标记检测[J]. 安徽农业大学学报,2010,37(3): 552 - 557.
[23] Liu Junli, Shen Hongxiang, Gao Xiahong,et al.A Primary Report of the Mutagenic Effects on Malus Ornamental Crabapple Induced by60Coγ-Irradiation [J]. Chinese Agricultural Science Bulletin, 2009, 25(6): 223 - 226.
[24] 张慧琴,谢 鸣,陈昆松,等.60Coγ辐照对草莓花药愈伤组织诱导和分化的影响[J]. 核农学报,2007,21(3): 229-231,267.
[25] 王小敏,吴文龙,张春红,等.60Co~γ辐照对黑莓组培苗的诱变效应及半致死剂量[J]. 经济林研究, 2011, 29(3): 35-39.
[26] 张 锐,王新建,吴翠云,等. 香梨60Co~γ辐射诱变效应初步研究[J]. 塔里木大学学报, 2006, 18(4): 5-9.
[27] 乐文全,李爱军,张海娥,等. 辐射对梨果实经济性状诱变效应的研究[C]//中国园艺学会梨分会第二届代表大会暨全国第五届梨科研、生产与产业化研讨会论文集. 上海:中国园艺学会,2007:74-77.
