不同种源无患子光合与蒸腾速率差异分析
2013-04-04孟德悦吴海勇刘光斌江春艳胡冬南
孟德悦,吴海勇,刘光斌,江春艳,刘 顺,胡冬南
(1.江西农业大学 园林与艺术学院,江西 南昌 330045;2.江西省靖安县林业局,江西 宜春 330600)
光合作用是植物将太阳能转换为化学能的过程,既是植物体内最重要的化学反应[1],又是植物生长发育的基础和植物体内碳素的重要来源[2],也是干物质积累的基础[3]和决定生产力高低的重要因素[4]。植物地上部分90%~95%的干物质来自于光合作用[5]。植物光合作用是一个对环境条件变化很敏感的生理过程[4],不但受环境因子的影响,而且受遗传基础的控制[6]。光合速率的日变化揭示光合作用的日进程,种源间光合特征的差异反映了不同种源光合能力的高低[7]。因此,对林木光合特性的研究,有助于了解林木生长发育规律,可为林木栽培抚育、引种、抗逆种子筛选和早期选择等提供理论依据[8]。
无患子Spindus mukorossiGaertn 系无患子科无患子属落叶乔木,在东南亚各国、我国的台湾省及淮河以南各省区均有分布[9]。其根系发达,耐干旱贫瘠,常见于低山、丘陵及石灰岩山地[10]。无患子树形优美,抗虫抗害能力强,为园林观赏、绿化造林的首选树种[11];无患子也是我国传统中药材[12],在开发生物洗涤剂及天然化妆品等方面应用广泛[13]。因无患子种仁含油量超过40%[14],故作为生物质原料树种的无患子受到了人们越来越多的关注[15]。目前,有关无患子育苗技术、开发利用等方面的研究报道较多,却未见到有关其种源差异方面的研究报道。为此,文中对无患子光合速率等生理指标进行了测定,在此基础上分析了不同种源无患子光合与蒸腾速率的差异性,以期为无患子的良种选育提供参考依据与技术支撑。
1 材料与方法
1.1 供试材料
供试无患子种子分别采自江西省的靖安县和九江市庐山区,2012年3月播种于江西农业大学科技园,测定时两地苗木的平均高度分别为130 cm和110 cm。
1.2 试验设计与测定方法
试验采用随机区组设计,共设计了3个处理,即种源(A)、树冠部位(B)、种源与树冠部位的交互作用(A×B)。试验因素A为种源(2 个),分别是靖安(A1)和九江(A2),每个种源重复3次;试验因素B为树冠部位(3 个),分别为上部(B1)、中部(B2)、基部(B3)3个水平;重复次数为6次(即6个观测时间点)。
分别从靖安县和庐山区这两个种源地的苗木中各选3株无患子作为标准木,采用L6400便携式光合测定仪,测定其树冠上部、中部、基部叶片(每片叶子重复测定5次)的净光合速率值和蒸腾速率值,从而得到其日变化曲线图。具体测定时间为2012年9月27日(天气晴好)的8:00 ~ 18:00,时间间隔为 2 h,即分别在 8:00、10:00、12:00、14:00、16:00、18:00 这 6 个时间点进行测定。
1.3 数据统计与分析
采用Excel2003和DPS v7.05版软件对文中的数据进行统计和分析。
利用DPS v7.05方差分析计算软件进行两因素有重复固定模型的方差分析和多重比较。
2 结果与分析
2.1 无患子叶片净光合速率的日变化曲线
2.1.1 不同种源无患子叶片净光合速率的日变化曲线
在不同测定时间点上测得的不同种源的无患子叶片的净光合速率值各不相同,其日变化曲线图如图1所示。从图1中可以看出,在6个时间点上测定的叶片净光合速率值,九江的无患子始终大于靖安的无患子,16:00以前两个种源的无患子其净光合速率值之间相差较大,但16:00和18:00测定的净光合速率值两者接近;两个种源的无患子叶片其净光合速率变化曲线总体上都呈现出早晚较低而中午12:00时最高的单峰抛物线,上午净光合速率随光照的加强、温度的升高而升高,到中午达到峰值后,下午又随光照强度和气温的降低而降低,故呈单峰曲线。

图1 不同种源无患子叶片净光合速率的日变化曲线Fig.1 Diurnal changes of Pn in S. mukorossi leaves from different provenances
2.1.2 无患子叶片净光合速率的差异性分析
不同时间点测定的靖安和九江2个种源的无患子苗木上、中、基部3个树冠部位叶片的净光合速率见表1。
根据表1的数据,对种源和树冠部位2个因素6次重复测定的净光合速率作方差分析,其中A因素有靖安、九江2个水平,B因素有上、中、基部3个水平,结果见表2。由表2可知,不同种源、交互作用(A×B)间存在极显著差异(p<0.01),不同树冠部位间存在显著差异(p<0.05)。

表1 不同时间测定的不同种源和树冠部位无患子叶片的净光合速率Table 1 Pn in S. mukorossi leaves at different determination time from different provenances and crown positions
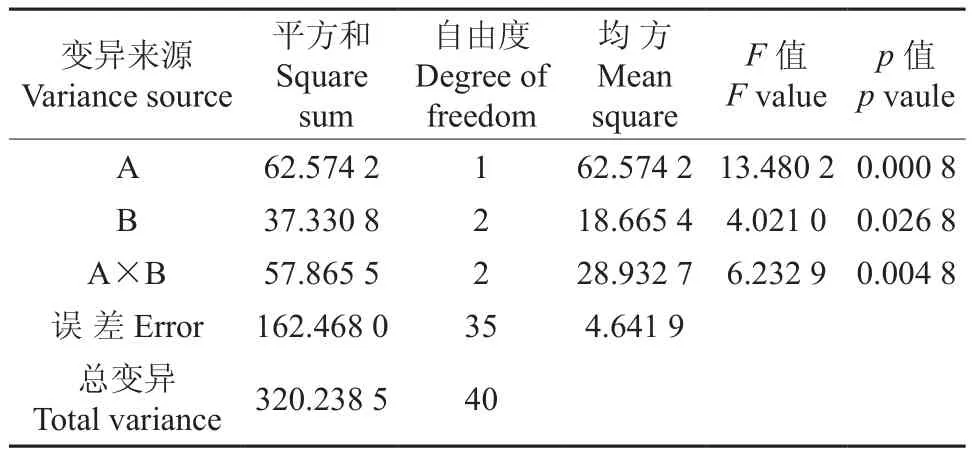
表2 不同种源和树冠部位无患子叶片净光合速率的方差分析结果Table 2 Variance analysis result of Pn in S. mukorossi leaves from different provenances and crown positions
为了进一步探究各因素间净光合速率的差异,对所得数据进行了多重比较分析,结果如表3所示。从表3中可以看出,2个种源无患子叶片的净光合速率达到了显著差异水平(p<0.05),九江无患子叶片的净光合速率显著大于靖安的叶片净光合速率。树冠上、中部叶片的净光合速率显著大于基部叶片。这是由于在生长发育过程中,受季节、光照历程、栽植密度、生态小环境等因素的影响,上、中部叶绿素含量相对较高造成的。九江中部叶片的净光合速率最高,显著大于上、基部与靖安3个部位叶片的净光合速率,其次为靖安上部叶片的净光合速率,而靖安基部叶片的净光合速率最低。这除了与外界环境、栽植密度等因素有关外,还与不同部位叶片的光能捕获能力与转化效率有关。

表3 不同处理无患子叶片净光合速率间的多重比较分析结果Table 3 Multiple comparison result of Pn in S. mukorossi leaves between different treatments
2.1.3 不同部位叶片净光合速率的日变化曲线
6个时间点观测到的同一种源不同部位叶片的净光合速率值各不相同。现据所测数据分别作出了靖安和九江的无患子叶片净光合速率值的日变化图,分别如图2、图3所示。

图2 靖安无患子不同部位叶片净光合速率的日变化曲线Fig.2 Diurnal changes of Pn in leaves at different positions of S. mukorossi crown from Jing’an
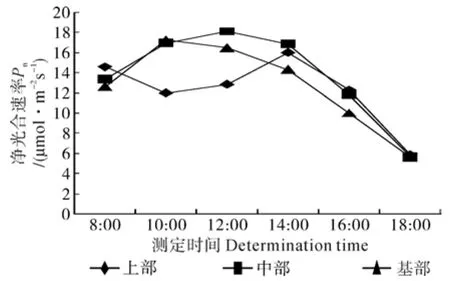
图3 九江无患子不同部位叶片净光合速率的日变化曲线Fig.3 Diurnal changes of Pn in leaves at different positions of S. mukorossi crown from Jiujiang
由图2可知,在6个测定时间点内,靖安的上、中部叶片净光合速率均高于基部叶片的净光合速率;上、基部叶片净光合速率的变化曲线呈现出单峰抛物线型,中部叶片呈现出双峰抛物线型,2个峰值分别出现在12:00和16:00,两峰值间的低谷值出现在14:00,且第1次峰值大于第2次峰值。“午休”现象的出现与否,与测定当天的光照和气温等影响因素有密切关系。
由图3可知,九江无患子中、基部叶片的净光合速率均呈先上升后下降的单峰曲线变化趋势,而上部叶片净光合速率则呈先下降后上升再下降的变化趋势。这与徐清乾等人[16]对四川桤木净光合速率的研究结果存在差异,导致这一现象的原因可能是无患子幼苗上部存在新发叶片,在叶片选择过程中存在一定差异;上部的叶片组织幼嫩,在仪器操作过程中可能会对叶片造成一定程度的损伤。
2.2 无患子叶片蒸腾速率的日变化曲线
2.2.1 不同种源无患子叶片蒸腾速率的日变化曲线
6个时间点观测到的不同种源无患子叶片的蒸腾速率值各不相同,其日变化曲线图如图4所示。从图4中可以看出,2个种源的无患子叶片的蒸腾速率都呈现出单峰型变化曲线,但九江的峰值出现在12:00,而靖安的峰值出现在14:00;九江无患子叶片的蒸腾速率始终大于靖安无患子叶片的蒸腾速率,这一点与两个种源无患子叶片净光合速率的日变化曲线一致。

图4 不同种源无患子叶片蒸腾速率的日变化曲线Fig.4 Diurnal changes of Tr in S. mukorossi leaves from different provenances
比较无患子叶片的净光合速率与蒸腾速率日变化曲线后可以发现,这2个指标具有相关性,净光合速率与蒸腾速率的峰值出现时间和变化规律基本一致。
2.2.2 不同种源无患子叶片蒸腾速率的差异性分析
不同时间点测定的靖安和九江2个种源的无患子苗木上、中、基部3个树冠部位叶片的蒸腾速率见表4。

表4 不同时间测定的不同种源和树冠部位无患子叶片的蒸腾速率Table 4 Tr in S. mukorossi leaves at different determination time from different provenances and crown positions
根据表4的数据,以种源和树冠部位2因素6重复作方差分析,结果见表5。由表5可知,两种种源间存在极显著差异(p<0.01),不同树冠部位间不存在显著差异,而交互作用A×B内各水平也达到了显著差异水平(p<0.05)。
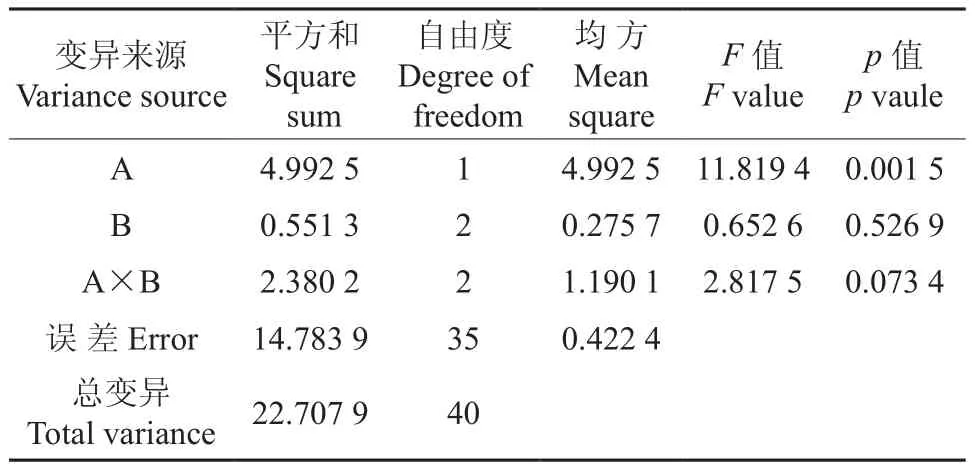
表5 不同种源和树冠部位无患子叶片蒸腾速率的方差分析结果Table 5 Variance analysis result of Tr in S. mukorossi leaves from different provenances and crown positions
为进一步分析各因素间蒸腾速率的差异性,对所得数据进行了多重比较,结果见表6。由表6可知:九江无患子叶片的蒸腾速率要显著大于靖安无患子叶片的蒸腾速率;九江中、基部叶片的蒸腾速率与上部叶片蒸腾速率差异不显著,但显著大于靖安中、基部叶片的蒸腾速率。

表6 不同处理无患子叶片蒸腾速率间的多重比较结果Table 6 Multiple comparison results of Tr in S. mukorossi leaves between different treatments
2.2.3 不同部位叶片蒸腾速率的日变化曲线
在6个时间点上测得的同一种源不同部位无患子叶片的蒸腾速率值各不相同。现据所测数据分别对靖安和九江的无患子叶片的蒸腾速率作了日变化规律图,分别如图5、图6所示。

图5 靖安无患子不同部位叶片蒸腾速率的日变化曲线Fig.5 Dinurnal changes of Tr in leaves at different positions of S. mukorossi crown from Jing’an

图6 九江无患子不同部位叶片蒸腾速率的日变化曲线Fig.6 Dinurnal changes of Tr in leaves at different positions of S. mukorossi crown from Jiujiang
由图5可知,在6个观测时间上,靖安上、中部叶片的蒸腾速率始终高于基部叶片的蒸腾速率,14:00~16:00上部叶片蒸腾速率开始低于中部叶片的蒸腾速率,并持续到18:00;3个部位叶片的蒸腾曲线均呈单峰抛物线,上、中部叶片的峰值出现在12:00,而基部叶片的峰值出现在14:00。
从图6中可以看出,九江3个部位叶片的蒸腾速率均呈现出形状不同的单峰抛物线,但中、基部的峰值出现在12:00,而上部叶片的峰值出现在14:00。这是叶片在不同时间受到光照、温度等因素的影响,在叶片自我调节能力[17]下,对水分蒸腾产生控制所致。
2.3 无患子叶片水分利用率的日变化曲线
水分利用率反映了植物利用水的效率,由净光合速率与蒸腾速率的比值求得。
2.3.1 不同种源叶片水分利用率的日变化
在不同时间点上测到的不同种源无患子叶片的水分利用率值各不相同,其日变化曲线图如图7所示。从图7中可以看出,靖安叶片水分利用率最低值出现在14:00,而九江叶片水分利用率最低值出现在 12:00,在 12:00 与 16:00 时,两个种源的水分利用率几乎重合,但2个种源的叶片水分利用率的变化规律基本一致,均呈先降低后升高的变化趋势。

图7 不同种源无患子叶片水分利用率的日变化曲线Fig.7 Diurnal changes of WUE in S. mukorossi leaves from different provenances
2.3.2 无患子叶片水分利用率的差异性分析
不同时间点测定的靖安和九江2个种源的无患子苗木上、中、基部3个树冠部位的水分利用率如表7。

表7 不同时间测定的不同种源和树冠部位无患子叶片的水分利用率Table 7 WUE in S. mukorossi leaves at different determination time from different provenances and crown positions
以种源、树冠部位2个因素的水分利用率作方差分析,结果如表8所示。从表8中可以看出,不同种源、树冠部位间的水分利用率差异不显著(p>0.05),且不同种源与树冠部位的交互作用A×B间也不存在显著差异(p>0.05)。说明试验立地的水分条件能够满足2个种源对无患子水分的要求,但是由于本试验所测定的水分利用率表征的是瞬时水分利用效率[18],且受当时净合速率、蒸腾速率的影响较大,因此仅代表测定时的水分利用情况。

表8 不同种源和树冠部位无患子叶片水分利用率的方差分析结果Table 8 Variance analysis result of WUE in S. mukorossi leaves from different provenances and crown positions
3 结论与讨论
通过对不同种源和不同树冠部位无患子叶片的净光合速率与蒸腾速率日变化的比较分析,结果表明:
(1)2个种源的无患子叶片的净光合速率和蒸腾速率均呈单峰抛物线趋势,且九江的均高于靖安。
(2)在2个种源的不同树冠部位叶片净光合速率与蒸腾速率日变化中,只有靖安中部叶片的净光合速率日变化出现双峰抛物线,峰值分别出现在 12:00 和 16:00,两峰间的低谷出现在 14:00,且第1次峰值大于第2次峰值,而九江上部叶片的净光合速率出现先下降后升高再下降的趋势,造成这种现象的原因是多方面的,其他均为单峰抛物线。“午休”现象不稳定的出现,这一点可能与测定当天的光照、温度和测定时间等因素有关,其原因尚待进一步开展研究。
(3)九江中部叶片、靖安上部叶片的净光合速率和蒸腾速率分别大于其他部位,说明九江中部叶片、靖安上部叶片的光合作用分别强于其他部位。
(4)不同种源、树冠部位及二者交互作用间,叶片净光合速率均达到显著差异水平,蒸腾速率除在树冠部位间差异不显著外,其他两者也达到显著差异水平,而水分利用率在这3个因素间均不存在显著差异。由于不同植物光合作用的大小不同,即使是同一种植物,因其基因型、品种、种源地的不同也会表现出光合作用的差异。
参考文献:
[1] 佘诚棋,程 鹏,季琳琳,等.油茶光合作用光响应曲线的拟合[J].经济林研究,2012,30(1):118-120.
[2] 韦铄星,文仕知,张 希,等.不同林龄四川桤木的光合生理特性分析[J].中南林业科技大学学报,2011,31(7):102-108.
[3] 姜成英,莫保儒,吴文俊,等.不同品种油橄榄的光合生理分析[J].经济林研究,2010,28(2):24-28.
[4] 高 超,闫文德,田大伦,等.杜仲光合速率日变化及其与环境因子的关系[J].中南林业科技大学学报, 2011, 31(5):100-104.
[5] Li D Y, Zhang Z A, Zheng D J,et al. Comparison of net photosynthetic rate in leaves of soybean with different yield levels [J]. Journal of Northeast Agricultural University, 2012,19(3): 14 - 19.
[6] 廖菊阳,闫文德,王光军,等.鹿角杜鹃光合日变化特性分析[J].中南林业科技大学学报,2011,(5):117-120.
[7] 杨瑞卿,楚秀丽,杨万霞,等.不同种源青钱柳苗期光合特征及变异分析[J].南京林业大学学报:自然科学版,2012, 36(4):41-45.
[8] 冯 健,王骞春,颜廷武,等.不同种源东部白松苗期光合作用研究[J].辽宁林业科技,2011,(2):5-7,60,62.
[9] 辜夕容.不同种源无患子的种子品质差异分析[J].西南大学学报(自然科学 版),2009,31(6):51-54.
[10] 徐启定.无患子的生态功效和利用价值[J].安徽林业,2010,(3):64.
[11] 黄 顺.优良的造景树-无患子[J].园林,2004,(9):42.
[12] 周爱红.无患子的用途及栽培技术[J].林业科学,2011,(7):213.
[13] 林文荣.无患子开发利用前景[J].林业勘察设计, 2007, (2):118-120.
[14] 王建章,吴子斌.无患子籽油成分分析与提取工艺研究[J].农业科学研究,2010,31(1):48-50.
[15] 闫 瑞,钱 春.新型生物质能源树种无患子在重庆的发展优势分析[J].南方农业,2012,6(6):103-105.
[16] 徐清乾,周小玲,许忠坤.四川桤木品系的光合和蒸腾速率差异及其生长分析[J].林业科技开发,2008,22(4):42-45.
[17] 宋宏伟,张江涛,王 晶,等.黄连木光合特性和水势变化规律的研究[J].安徽农业科学.2007,35(31):9924-9926.
[18] 张凤良,张方秋,段安安,等.不同种源红锥光合特性与水分利用效率变异分析[J].西南林业大学学报, 2012,32(4):12-16.
