2007—2008年春夏季长江口水域浮游动物生态分布特征研究
2013-03-20刘守海项凌云刘材材王金辉
刘守海,项凌云,刘材材,王金辉
(国家海洋局东海环境监测中心 国家海洋局海洋赤潮灾害立体监测技术与应用重点实验室,上海 200137)
浮游生物是指在水流运动的作用下,被动地漂浮在水层中的生物群。它们的共同特点是缺乏发达的运动器官,运动能力薄弱或完全没有运动能力,只能随水流移动(沈国英等,2002)。浮游动物在整个海洋生态系统中起着非常重要的调控作用,其动态变化控制着初级生产力的节律、规模和归宿,也直接影响到整个生态系统的变化(徐兆礼等,2003;Zhang et al,2010)。
长江口位于河流与海洋的交汇处,是二者相互作用的复杂区域,其营养物质丰富,生物生产力大,生态环境多变。长江口因其独特的河口环境条件,其浮游动物的生态研究一直是研究的热点。有关长江口及邻近水域浮游动物的研究早有报道(陈亚瞿 等,1985,1995a,1995b;朱启琴,1988;徐兆礼 等,1995,1999,2005a),但较少涉及到整个河口范围的调查(纪焕红等,2006;郭沛涌等,2003,2008a;徐韧等,2009),且极少有连续几年同地点的资料比较。根据2007—2008年连续两年在5月、8月在长江口水域的定点调查资料,对该海域浮游动物群落的分布特征及其影响因子进行了研究,以期为长江口水域浮游动物的长期变化研究提供基础资料。
1 材料与方法
2007年到2008年分别于5月和8月对长江口区(121°15′-123°10′E,30°40′-31°45′N) 海域进行监测调查,共布设20 个站(图1)。根据参考文献(纪焕红 等,2006;徐韧 等,2009;王金辉等,1995;郭沛涌等,2008b),可将本调查水域分为3 个区域:长江口河口水域(1-4 号站)、长江口咸淡交错水域(5-10 号站) 和长江口外近海水域(11-20 号站)。
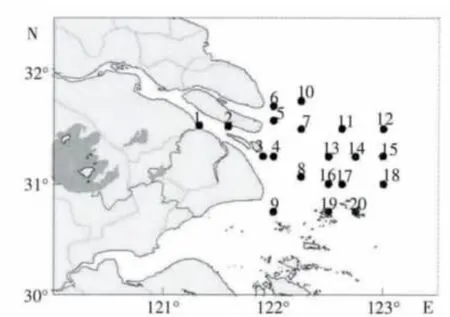
图1 采样站位
浮游动物样品采集及室内处理等均按《海洋调查规范》 (国家质量监督检验检疫总局,2007) 进行。用浅水Ⅰ型浮游生物网 (口径50 cm、筛绢CQ14、孔径0. 505 mm) 由底至表层垂直拖网采集,所获样品均经5%福尔马林溶液固定,样品种类鉴定到种,并计数称重,丰度单位为ind/m3,生物量单位为mg/m3。
优势度(Y) 和Shannon-Weaver 生物多样度指数(H′) 计算公式见参考文献(徐兆礼 等,1995):
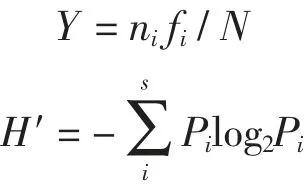
式中:ni为第i 种的丰度;fi是该种在各站位中出现的频率;N 为浮游动物总丰度;Pi表示第i 种的丰度(ni) 与总丰度(N) 的比值;S 为样品中总种数。取优势度Y ≥0.02(徐兆礼等,1989) 的浮游动物种类为优势种。
数据相关分析采用SPSS16.0 统计软件包Pearson 相关分析处理,其余数据处理利用Office 2007、SPSS 16.0 统计软件进行。
2 结果
2.1 浮游动物生物量变化及平面分布
调查水域浮游动物生物量值分布格局呈近岸向外海递增趋势(图2),即长江口河口水域(站位1号-4 号) <长江口咸淡交错水域(5 号-10 号站位)<长江口外近海水域(11 号-20 号站位);8月份生物量大于5月份生物量。
2007年5月生物量平均值为319.48 mg/m3,较高值位于口外近海水域的东部,即15 号站附近,为1 066.38 mg/m3。2008年5月生物量平均值为399.01 mg/m3,高值区位置位于口外近海中部水域,即122°30′E 附近,生物量值为2 283.57 mg/m3。
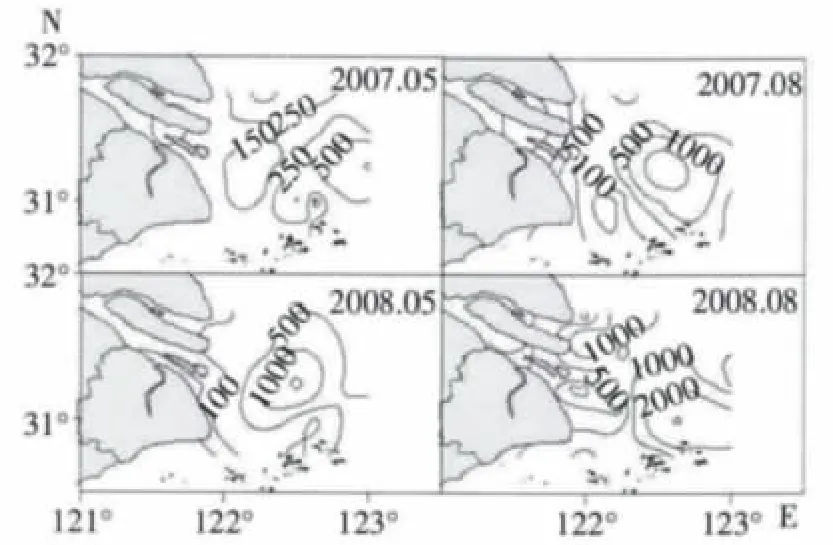
图2 浮游动物生物量平面分布(mg·m-3)
2007年8月生物量平均值为830.23 mg/m3,高值区几乎包括了河口水域和整个口外近海中部,即122°30′-122°50′E 附近水域。2008年8月生物量平均值为1 159.07 mg/m3,高值区位于口外近海水域的东南部水域,生物量高达5 421.77 mg/m3。
2.2 优势种及平面分布
中华哲水蚤(Calanus sinicus) 作为4 次采样均出现的种类,是整个调查水域的优势种(表1),尤其是口外近海水域的主要优势种。2007年和2008年5月都出现的优势种为虫肢歪水蚤(Tortanus vermiculus) 和真刺唇角水蚤(Labidocera euchaeta),两者多分布于河口水域以及咸淡交错水域,即122°30′以西水域。8月都出现的优势种为背针胸刺水蚤(Centropages dorsispinatus)、太平洋纺锤水蚤(Acartia pacifica) 和肥胖箭虫(Sagitta enflata);前二者分布于河口水域和咸淡交错水域,肥胖箭虫多分布于口外近海水域。

表1 不同年份不同季节优势种优势度
综合比较主要浮游动物种类的优势度、平均丰度及出现率大小,分析不同水域浮游动物的优势种变化(表2)。从表2 可知:除2008年8月外,中华哲水蚤均为优势种,且各区域均有分布。在河口水域,5月以华哲水蚤(Sinocalanus sinensis)、虫肢歪水蚤为主;8月以太平洋纺锤和背针胸刺水蚤为主;咸淡交错水域,5月以虫肢歪水蚤、真刺唇角水蚤为主,8月以背针胸刺水蚤、太平洋纺锤水蚤真刺唇角水蚤和虫肢歪水蚤为主;口外近海水域外,5月以中华哲水蚤为主,8月以中华哲水蚤、背针胸刺水蚤和肥胖箭虫为主,且种类数较多。
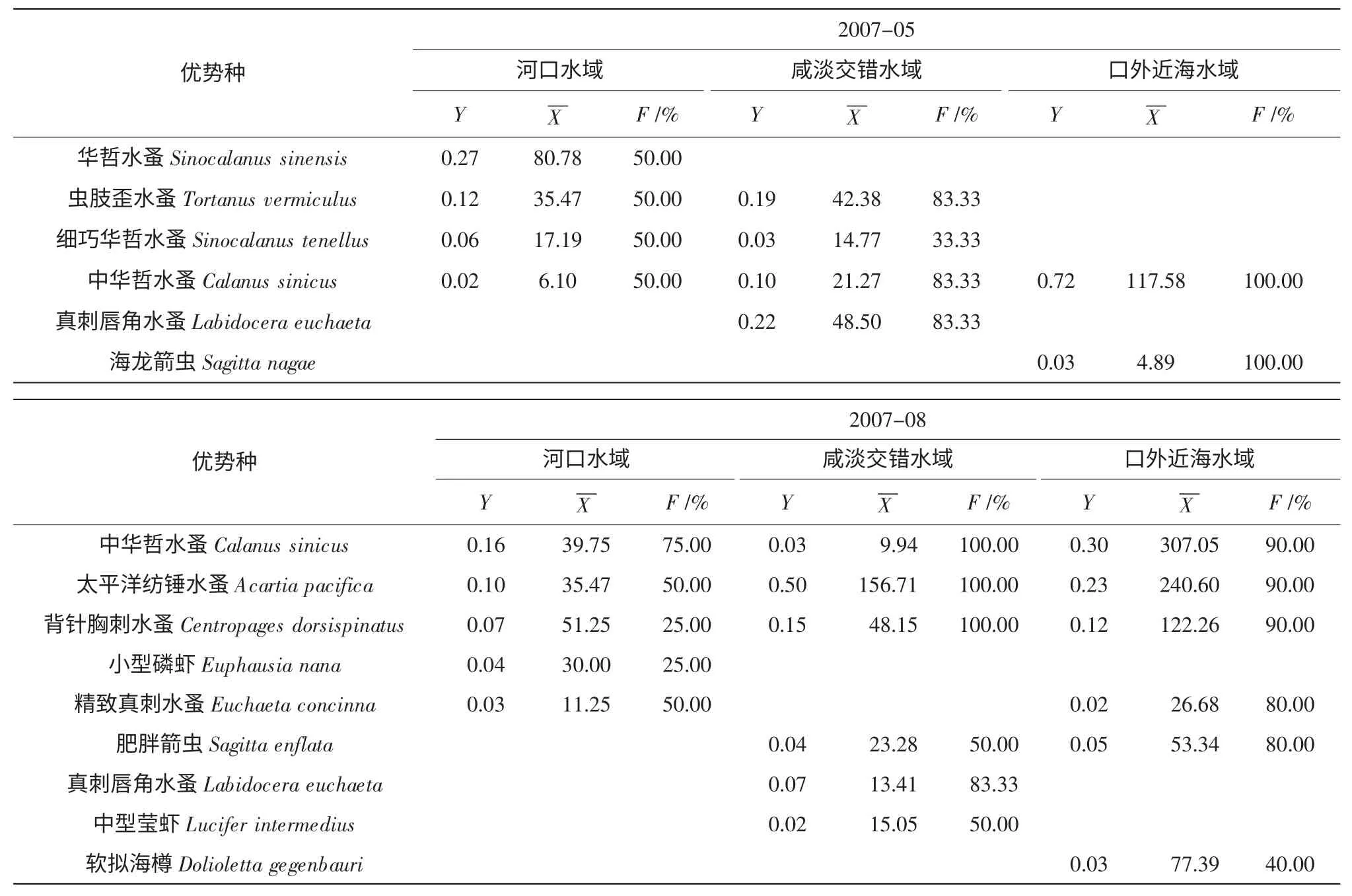
表2 不同水域优势种优势度及平均丰度
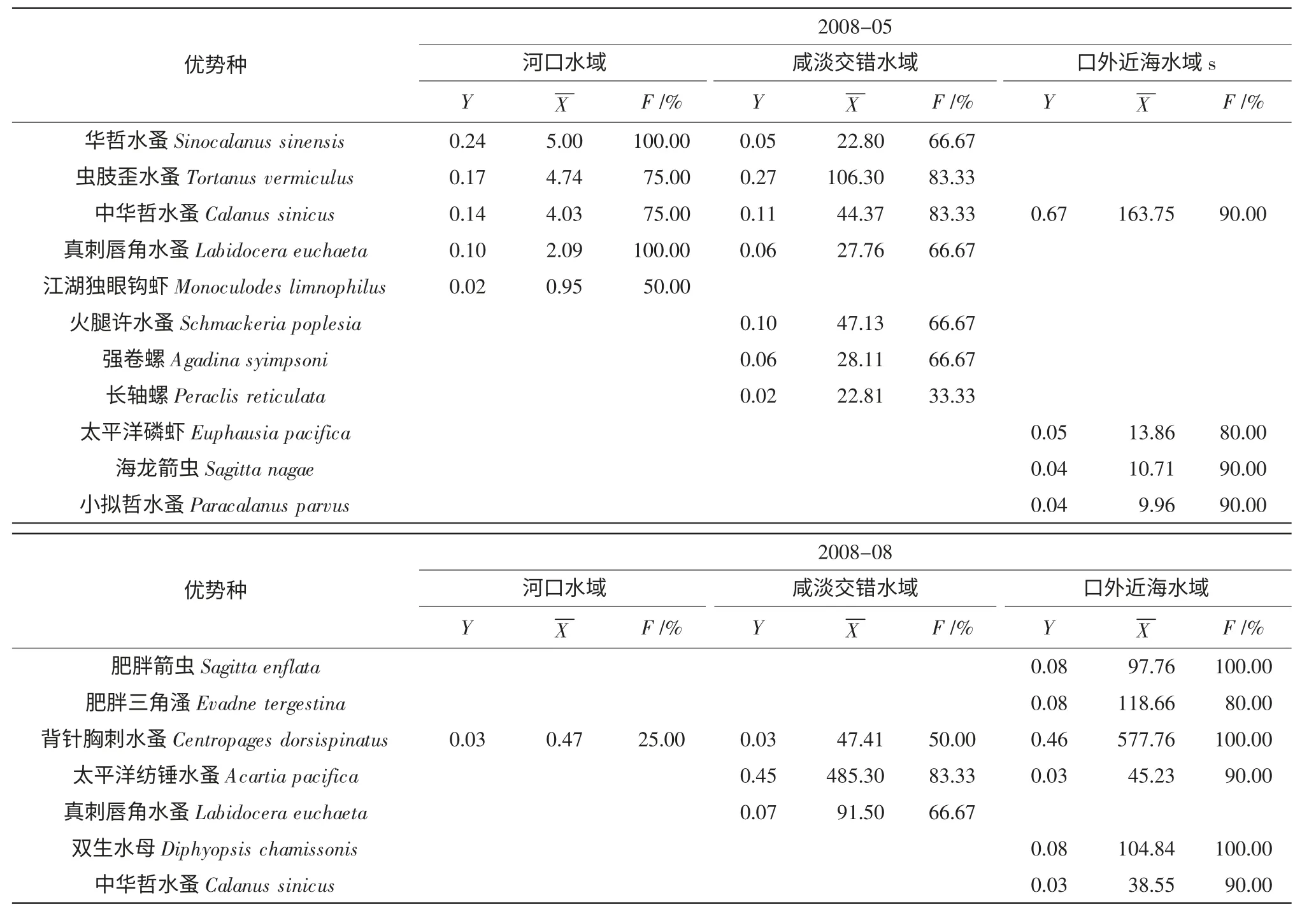
Y:优势度;X:平均丰度;F:出现频率
2.3 种类数和物种多样性的平面分布
调查水域浮游动物种类数分布格局呈近岸向外海递增趋势(图3),即长江口河口水域(站位1号-4 号) <长江口咸淡交错水域(5 号-10 号站位)<长江口外近海水域(11 号-20 号站位)。8月份总的种类数大于5月份的种类数。
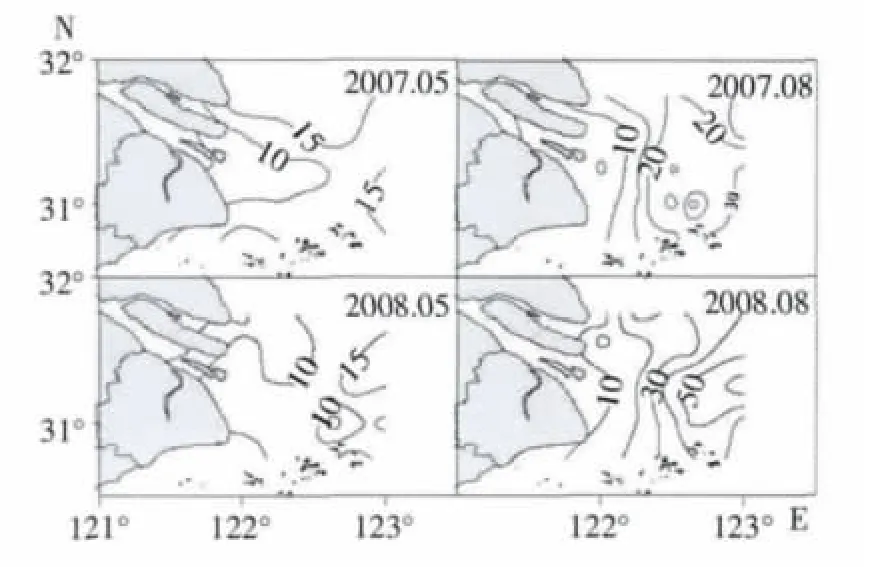
图3 浮游动物种类数平面分布
2007年5月种类数较高值位于口外近海水域和咸淡交错水域的靠北部分水域以及口外近海水域的东南部水域,即6 号站、10 号站11 号站之间的水域以及18 号站附近水域。2008年5月高值区位置位于口外近海水域,位于122°45′-123°E 附近。
2007年8月种类数高值区近乎包括了整个口外近海的调查水域,位于122°30′E 以东水域。与2007年相比,2008年8月种类数的高值较2007年8月更高,15 号站的种类数达65 种;但高值区的范围更集中,位于在口外近海水域的中部。
5月多样性指数分布不规律。2007年5月,最大值区域位于长江口咸淡交错水域的靠北支水域,均值为1.93;河口水域的多样性指数值最低。2008年5月,最大值区域位于长江口河口水域和长江口咸淡交错水域交界水域,即121°55′-122°15′E 水域;外近海水域的多样性指数值最低。
8月多样性指数均值分布格局大致为:长江口河口水域<长江口咸淡交错水域<长江口外近海水域(见图4)。
2.4 浮游动物与环境因素的关系
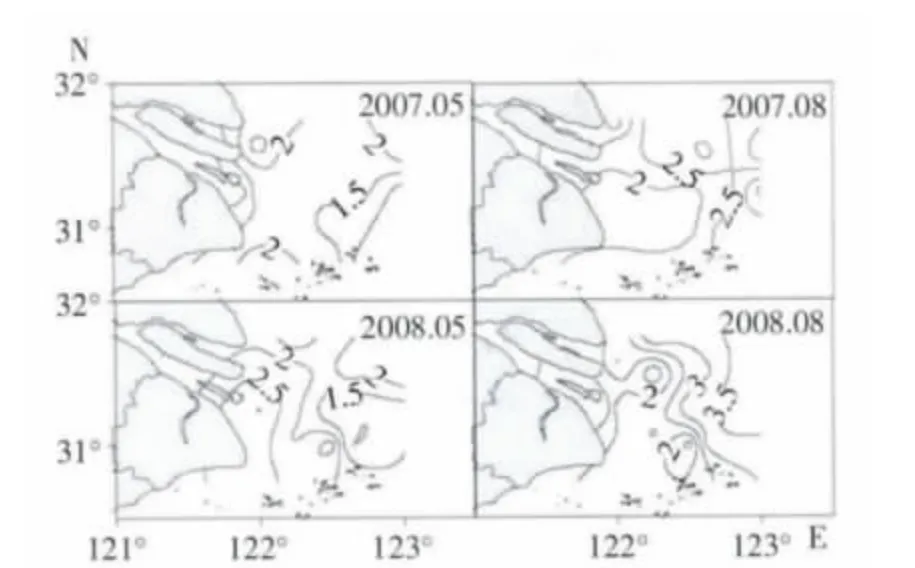
图4 浮游动物多样性指数平面分布
浮游动物的分布与温度、盐度等因子有关,其相关性分析的结果如表3 所示。2007年种类数变化与表层盐度呈线性正相关,2008年8月的种类数与底层水温呈线性负相关。2007年和2008年5月的多样性指数与中华哲水蚤丰度呈负相关。多样性指数还与底层水温、盐度等有关。
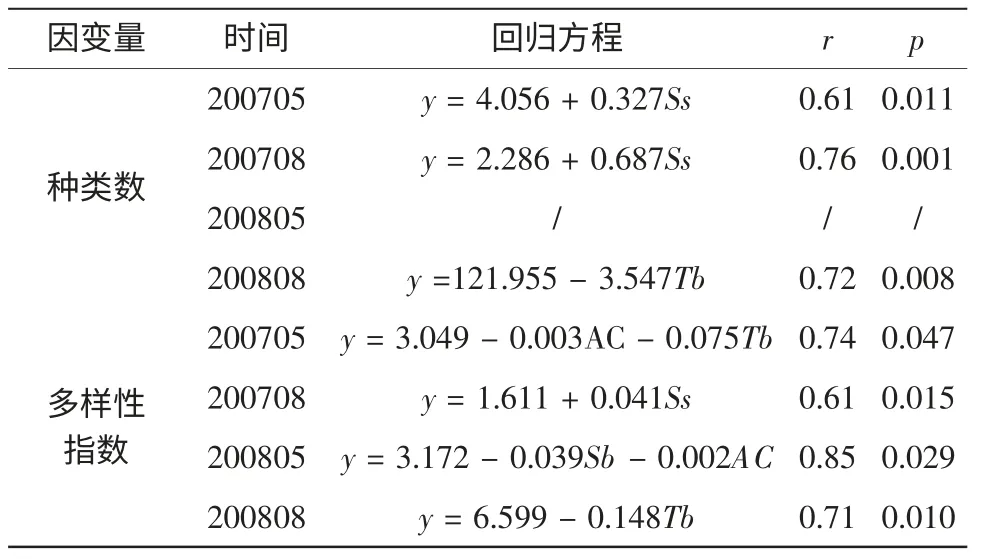
表3 浮游动物生态特征与环境因子的相关性分析
3 讨论
3.1 生物量变化与优势种类的关系
桡足类是调查水域的主要优势种类(表1),与之前的研究结果一致(朱启琴,1988;陈亚瞿等,1995a,1995b;徐兆礼等,1995,1999,2005a;纪焕红等,2006;郭沛涌等,2003)。近岸种中华哲水蚤是本次调查最重要的优势种,是2007年5月、8月和2008年5月的绝对优势种(表1);2008年8月中华哲水蚤优势度下降明显,是由于河口水域和咸淡交错水域出现了较高密度的背针胸刺水蚤、太平洋纺锤水蚤。此外,咸淡交错水域5月的主要优势种是虫肢歪水蚤和真刺唇角水蚤,8月主要是太平洋纺锤水蚤(表2)。徐韧等(2009) 的研究结果认为河口种真刺唇角水蚤是长江口咸淡交错水域5月份和8月份最主要的优势种,这一结果与本研究的结果有较大不同,这可能与浮游动物以及长江径流量的年际变化有关。
浮游动物生物量的变化与优势种丰度变化密切相关。本研究的生物量高值及较高值都出现在口外近海水域及咸淡交错水域与口外近海水域交界水域(图2),其中中华哲水蚤是最主要的优势种(表1),且中华哲水蚤个体较大,因此中华哲水蚤对生物量的贡献较高。中华哲水蚤成为优势种的水域标志着外海高盐水入侵的位置(徐兆礼等,2005c),说明调查水域可能受到外海水的影响。
3.2 种类数与多样性特征的关系
春夏季调查水域浮游动物种类数分布格局总的来说,呈近岸向外海递增趋势(图3)。8月份总的种类数大于5月份的种类数,8月份的多样性特征的分布与8月份种类数的分布基本一致,高值区在口外近海水域,低值区出现在河口水域;而5月份种类数和多样性指数的分布有所差异。这说明种类数越多对多样性指数的影响越大。以浮游动物种类数(x) 为自变量,H′值为因变量进行线性模型拟合,表达式如下:
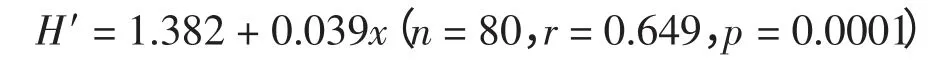
相关分析结果表明,种类数和H′值具有非常明显的相关关系,种类数的多少是决定调查水域大多数站位H′高低的主要因素。
3.3 浮游动物与环境的关系
浮游动物的分布与水温、盐度、水系、径流、浮游植物和叶绿素a 等因子密切相关(徐兆礼等,2003;陈洪举等,2006,2009)。盐度、水温是影响长江口及邻近水域浮游动物群落结构的主要环境因子(徐兆礼 等,2003,2005b;陈洪举 等,2006,2009;刘光兴等,2007),本文重点讨论调查水域盐度、水温对浮游动物分布的影响。
盐度是影响浮游动物群落特征重要的环境因子(Gouveia et al,1999;Rvetto et al,1999;Dauvin,2002;徐兆礼,2005b;刘光兴等,2007)。如表3所示,2007年5月表层盐度与种类数呈正相关;2007年8月表层盐度与种类数呈显著正相关,与多样性指数呈正相关(表3)。在长江口河口水域其盐度小于2,该水域浮游动物的种类数小于10。盐度高值区域出现在口外近海水域,盐度接近30,同时该水域浮游动物种类数亦是最多(图3)。在长江口水域,盐度平面分布呈向外海递增,种类数的分布规律亦是如此(图3)。多样性指数与种类数的分布规律基本一致,外海大于近岸(图4)。因此盐度影响种类数的分布,也影响多样性指数的变化。
此外,种类数、种类分布与盐度的分布规律关系密切。在河口水域的1,2 号站位中出现了淡水种类如广布中剑水蚤(Mesocyclops leuckarti)、英勇剑水蚤(Cyclops strenuus) 等;在咸淡交错水域以虫肢歪水蚤、火腿许水蚤、华哲水蚤等河口半咸水种类为优势种;高盐种类的肥胖箭虫出现在长江口外近海水域的14、17、18 号站。黄加祺等(1997) 对福建省南日岛海域浮游动物分布进行研究时还发现,在盐度高的季节,浮游动物出现的种类数也多,种类数的季节分布曲线与盐度的季节变化曲线比较吻合。此外,种类数与盐度的平面分布规律也比较一致。本文结果与其基本一致。
温度是影响浮游动物分布的一个重要因素(Dauvin,2002;Gouveia et al,1999;Sommer,1989;黄加祺等,1991,2000)。通常认为浮游动物的许多种类在12~17℃生长良好,在最适温度条件下会大量繁殖,温度较低或较高都会限制浮游动物的生长(徐兆礼等,1989)。本研究表明,2008年8月底层温度与种类数呈显著负相关,与多样性指数呈负相关;2007年5月底层温度与多样性指数呈负相关(见表3)。从温度变化看,温度分布由河口近岸向口外近海水域降低。而浮游动物种类数分布格局呈近岸向外海递增趋势(图3),浮游动物生物量分布是外海大于近岸(图2)。
陈洪举等(2009) 对2006年夏季长江口及其邻近水域浮游动物的群落结构研究表明,在单因子分析中,浮游动物丰度与底层温度的相关系数为0.308,与表层温度的相关系数为0.251。浮游动物分布与温度呈正相关。这一结果与本研究的结果有较大不同,这可能与研究的季节不一致有关。另外更重要的是本研究的区域更靠近长江河口区域,包括了种类较少但温度偏高的长江口河口水域,以及大部分咸淡交错水域。
4 结论
(1) 调查水域浮游动物生物量值8月份大于5月份生物量,分布格局呈近岸向外海递增趋势。
(2) 在河口水域及咸淡交错水域,5月的主要优势种为虫肢歪水蚤和真刺唇角水蚤,8月主要优势种为背针胸刺水蚤和太平洋纺锤水蚤。
(3) 调查水域浮游动物种类数和多样性指数分布格局呈近岸向外海递增趋势,种类数的多少是决定调查水域大多数站位H′高低的主要因素。
(4) 盐度、温度及主要优势种中华哲水蚤是影响长江口浮游动物分布的重要因子。
Dauvin J C,2002. Environmental control of mesozooplankton community structure in the Seine estuary.Oceanal Acta,25(1):13-22.
Gouveia I,Miguel C,et al,1999. Plankton of Guadiana:seasonal distribution and relation with environmental parameters-Preliminary results.Rev Biol Lisboa,17(1-4):179-192.
Rvetto G,Gaudy R,Pagano M,1999.Influence of salinity on the distribution of Acartia tonsa.J Exp Mar Biol Ecol,239(1):33-45.
Sommer U E,1989. Plankton Ecology,succession in plankton communities.Berlin:Spring-Verlag,253-296.
Zhang D j,Li S j,Guo D h,2010. Impacts of global warming on marine zooplankton.Marine Science Bulletin,12(2):15-25.
陈洪举,刘光兴,2009.2006年夏季长江口及其邻近水域浮游动物的群落结构.北京师范大学学报(自然科学版),45(4):393-397.
陈亚瞿,徐兆礼,王云龙,等,1995a.长江口河口锋区浮游动物生态研究Ⅰ.生物量及优势种的平面分布.中国水产科学,2(1):49-58.
陈亚瞿,徐兆礼,王云龙,等,1995b.长江口河口锋区浮游动物生态研究Ⅱ.种类组成、群落结构、水系指示种.中国水产科学,2(1):59-63.
陈亚瞿,郑国兴,朱启琴,1985.长江口区浮游动物初步研究.东海海洋,3(3):53-61.
郭沛勇,沈焕庭,刘阿成,等,2008b.长江河口中小型浮游动物数量分布、变动及主要影响因素.生态学报,28(8):3517-3526.
郭沛涌,沈焕庭,刘阿成,等,2003.长江河口浮游动物的种类组成、群落结构及多样性.生态学报,23(5):892-900.
郭沛涌,沈焕庭,刘阿成,等,2008a.长江口桡足类数量分布与变动.生态学报,28(9):4259-4267.
国家质量监督检验检疫总局,2007.海洋调查规范,第6 部分:海洋生态调查.北京:中国标准出版社,34-38.
黄加祺,洪幼环,朱长寿,等,1997.福建南日岛浮游动物的分布.东海海洋,15(4):46-53.
黄加祺,李少菁,陈亚萍,1991.福建罗源湾浮游桡足类的分布.台湾海峡,10(1):46-51.
黄加祺,李少菁,侯红兵,2000.福建罗源湾浮游桡足类群落特征.海洋科学,24(6):1-4.
纪焕红,叶属峰,2006.长江口浮游动物生态分布特征及其与环境的关系.海洋科学,30(6):23-30.
刘光兴,陈洪举,朱延忠,等,2007.三峡工程一期蓄水后长江口及其邻近水域浮游动物的群落结构. 中国海洋大学学报,37(5):789-794.
沈国英,施并章,2002.海洋生态学.北京:科学出版社,37.
王金辉,黄秀清,刘阿成,等,1995.长江口及邻近水域的生物多样性变化趋势分析.海洋通报,23(1):32-39.
徐韧,李亿红,李志恩,等,2009.长江口不同水域浮游动物数量特征比较.生态学报,29(4):1688-1696.
徐兆礼,2005a.长江口北支水域浮游动物的研究.应用生态学报,16(7):1341-1345.
徐兆礼,2005b.长江口邻近水域浮游动物群落特征及变动趋势.生态学杂志,4(7):780-784.
徐兆礼,陈亚瞿,1989.东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系.生态学杂志,8(4):13-15.
徐兆礼,洪波,朱明远,等,2003.东海赤潮高发区春季浮游动物生态特征的研究.应用生态学报,14(7):1081-1085.
徐兆礼,沈新强,马胜伟,2005c.春、夏季长江口邻近水域浮游动物优势种的生态特征.海洋科学,29(12):13-19.
徐兆礼,王云龙,白雪梅,等,1999.长江口浮游动物生态学研究.中国水产科学,6(5)增刊:55-58.
徐兆礼,王云龙,陈亚瞿,等,1995.长江口最大浑浊带浮游动物的生态研究.中国水产科学,2(1):39-48.
朱启琴,1988.长江口区浮游动物生态调查报告.水产学报,12(2):111-123.
