桑螟发生规律及防治技术的研究进展
2013-03-15赵叙华白锡川施国方
赵叙华,白锡川,施国方
(1.湖州市农业科学研究院,浙江湖州 313000;2.吴兴区农林技术推广服务中心,浙江湖州 313000)
桑螟发生规律及防治技术的研究进展
赵叙华1,白锡川2,施国方1
(1.湖州市农业科学研究院,浙江湖州 313000;2.吴兴区农林技术推广服务中心,浙江湖州 313000)
桑螟分布广、繁殖力和扩散能力强,防治难度大,已逐渐上升为桑园主要害虫,近年来已成为桑保界研究的热门课题。本文叙述了目前全国各地对桑螟发生规律、生活习性、防治技术等方面研究的一些概况,为桑螟进一步研究提供参考。
桑螟;发生规律;防治技术
桑螟[Diaphania pyloalis(Walker)]属鳞翅目螟蛾科,谷名白蚰、油虫、卷叶虫、青虫等[1]。除新疆外国内蚕区均有发生,江苏、浙江、广东、安徽、江西、四川、湖南、湖北、广西、重庆尤为严重。国外分布于东南亚、日本、印度等国。桑螟寄主植物单一,至目前除危害桑树外还未发现在其它作物上危害。
桑螟在20世纪80年代以前一直不是桑树主要害虫,80年代后在江、浙、渝一带成灾现象渐渐增多。1989年重庆北碚蚕区第3代桑螟桑树新梢危害率达67.6%,叶危害率46%[2];进入90年后,随着高效农药在桑园中推广使用,桑蟥、野蚕、桑尺蠖等桑园主要害虫被控制,在全国主要蚕区桑螟上升为一类主要害虫,多次出现桑螟严重危害,影响秋蚕正常生产。桑螟频繁暴发,不但严重影响蚕茧生产,而且由于桑螟的微粒子病与家蚕有互感性,诱发了蚕种生产微粒子病流行,上世纪90年代中后期,在江浙一带桑螟连续大发生。导致家蚕微粒子病大流行,蚕种淘汰率严重年份达到12.4%,蚕种带毒率超过80%,桑螟的发生严重影响到蚕种生产安全。
随着大量农药在桑园中使用,桑螟危害没有下降,反而有进一步加重之势。桑螟目前已经成为我国蚕区优势害虫,成为影响我国蚕桑生产安全的重大问题。对桑螟发生规律及防治方法研究成为全国桑保界一个重要课题,研究发表的文章不断增多。为了促进该领域研究工作的深入系统开展,本文简要综述有关桑螟发生规律及防治的研究进展,并针对该领域研究工作存在的一些问题,提出改进意见和建议。
1 生活习性
桑螟年发生代数自北向南为3~10代。在山东或山东附近年发生3代,少数年份4代;浙江、江苏、四川、安徽等地区年发生5代,少数年份有4代或6代发生[3,7,12];江西、贵州等地区年发生5~6代[13~14];福建等地区年发生7代[15];广西年发生8~10代[8]。以幼虫越冬,越冬盛期为11月。桑螟进入越冬状态是由短期光照和低温引起的兼性滞育,3、4龄虫期是光感应的重要阶段,完全滞育与非滞育,只有在一定的连续光照条件下才能产生。在温度24.6℃,纬度30°53′,桑螟的临界光周期为13 h 40min。解除滞育与春期有效积温有密切关系,早春气温回升快,桑螟发生也相应提前,一代桑螟出现迟早对全年桑螟代数多少密切关系[16]。桑螟越冬时间在不同地区有些差异,在太湖流域每年9月底大多数桑螟幼虫进入5龄期开始越冬,到10月中下旬越冬完毕[3];广西一带在10月虫口密度仍较大,危害严重,11月中旬开始,桑螟以老熟幼虫越冬[17]。桑螟越冬场所较多,在桑树上老熟幼虫在树干的缝隙、蛀孔、枯枝落叶和根际表土中越冬;被养蚕采叶带进蚕室的幼虫在建筑物墙缝、墙与墙和墙与楼板等连结处、室内堆放物等场所结白茧越冬,越冬时间长达250 d左右。越冬代桑螟幼虫羽化有二个特点,一是个体间羽化时间差异很大,前后可相差30多d,且羽化时间的迟早与越冬时间迟早和越冬场所关系不密切;二是越冬幼虫群体在羽化过程中峰值不明显[17]。所以桑螟有世代重叠现象,特别是桑螟大发生年,世代重叠尤为严重。
桑螟成虫羽化在晨3~9时,交尾后数h至24 h开始产卵,以第1~2 d产卵最多,第1 d产卵量占总卵数50%,第2 d占总卵量25%左右;产卵时间以晚上18~24时最盛;中午不产卵。产卵叶位,季节不同差异较大,春季产卵于桑枝新梢顶部1~9叶为多,秋季产卵于枝条中下部叶为多,但桑条顶端的芽、苞、新嫩叶产卵数多于其他部位叶[18]。
桑螟成虫善于飞行,在离桑园较远的城市高楼上也能引诱到桑螟蛾子;新开辟蚕区,桑树上最先出现的鳞翅目害虫一般都是桑螟。远离蚕区城市中间零星栽几株桑树也能出现桑螟严重危害。所以桑螟有远距离迁移能力。
桑螟卵小壳薄,在调查时非专业人员很难发现。遇高温干旱,卵易干瘪。卵孵以白天为主,孵化率在75%~95%左右,在气温低于16℃高于35℃时卵孵化率很差;秋季桑叶茂盛,常规性干旱对桑螟卵孵影响不大,如遇多雾天,孵化率可达到95%以上。卵在不同温度下历期分别是:21℃时为7.5 d左右,25℃时为6 d左右,28℃时为5 d左右,30℃时为4 d左右[20]。
桑螟幼虫初孵时很小,不细看很难发现。初孵幼虫居叶背分叉处,食害下表皮和叶肉。3龄后幼虫吐丝折叶或叠叶,潜伏其中进行取食,常见数头一起危害。一叶食光再移食其他叶,全叶食尽,则吐丝下垂,随风飘至其他株,或沿枝干下爬,向邻株迁移。一般经5次脱皮后老熟化蛹。各龄幼虫经过随季节气温变化差异较大,在太湖流域各龄幼虫经过如下表1[20]。
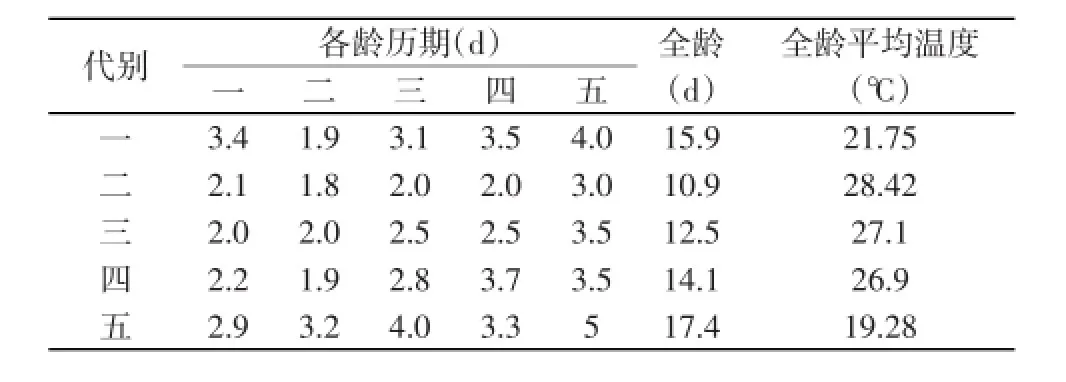
表1 桑螟各代幼虫历期Table 1 The period of each larval generation of mulberry moth
越冬桑螟幼虫化蛹时间受翌年早春有效积温控制,不同地区差异较大。在太湖流域一般在翌年4月中旬开始陆续化蛹,蛹期14~17 d左右,最长达到25 d,4月底能见到羽化初蛾,5月中下旬为羽化盛期。在广西地区2月中旬开始陆续化蛹,2月底、3月初陆续羽化。蛹期经过随温度变化也有些差异,当日平均温度在21℃时为14~15 d,24℃时为9~10 d,26℃时为8~9 d,28℃时为7 d,30℃时为7~8 d[3,8,9]。
2 发生规律
2.1 虫源基数
桑螟幼虫越冬基数与翌年早春一代桑螟发生量大小关系密切,上一年越冬代桑螟发生量大,翌年桑螟发生就重。但第一代桑螟发生量与以后各代桑螟发生量关系不密切。桑螟在自然状态下发生数量的变化幅度较大,前代桑螟数量多少与下一代桑螟数量多少不完全相对应。但桑螟暴发,必定有一个虫源基数。比如1997~1998年,在太湖流域2~3代桑螟基数很低,每公顷虫量在15000条以下,四代桑螟发生量得到了一些恢复,每公顷达到了21480条和19500条,五代桑螟大暴发,每公顷虫量达到600000条和570135条,桑叶损失率达到15%。浙江湖州地区,1999年3代桑螟每公顷虫量达到159960条,第四代桑螟每hm2虫量相应为339360条,桑叶损失率达10%;2001年四代桑螟每hm2虫量158400条,五代桑螟每hm2虫量相应为525600条,桑叶损失率达到11.2%[7,23]。
2.2 气候条件
桑螟卵的发育起点温度为12.41±4.13℃,有效积温63.14±8.68日度;幼虫的发育起点温度为11.12± 0.55℃,有效积温187.36±6.48日度;蛹的发育起点温度为13.40±1.11℃,有效积温94.27±8.40日度;成虫的有效积温为14.60±2.66℃,有效积温54.61±12.32日度;一世代的发育起点温度为12.95±0.93℃,有效积温为382.11±25.67日度[24]。
桑螟卵小、壳薄,易受气候影响,在多雾和多湿环境下卵孵化率高,相对湿度达到70%~80%时,卵孵化率可达到95%以上,甚至达到100%。农谚说“雾露多,生油虫”[1]。但遇高温干旱和强光易干瘪,孵化率大幅下降,严重时下降到50%以下。所以,正常年份,在大田中6~7月份桑螟发生数量是全年最低。根据浙江湖州地区调查,第一代桑螟幼虫基本上每公顷在22500条以上,第2~第3代(6~7月)每hm2在15000条左右,4~5代每公顷基本上150000条以上[7]。桑螟幼虫在多雨多湿环境中生存率降低,遇高温干旱能加剧桑螟危害程度。在太湖流域,秋季8~9月,桑园长势茂盛,内部小环境湿度较高,而大气气候比较干燥,非常适合桑螟繁育,所以4~5代极易暴发成灾。因5代桑螟在太湖流域极易成灾,浙江湖州地区应用生物统计方法及分析各气象要素对5代桑螟进行重点观察分析其影响。8月上旬高温多湿、8月中旬高温干燥对5代桑螟发生量有明显促进作用;9月上旬多湿和适度低温、9月中旬干燥和适度高温有利于5代桑螟数量增加[27]。多湿环境有利于桑螟卵的生存和孵化,干燥环境有利于桑螟幼虫和蛹的生存。在桑螟羽化盛期或幼虫孵化初期,遭遇连续大雨,下一代桑螟虫量将大幅度下降,或危害高峰将延迟。如浙北地区在2012年9月上中旬,正是4代桑螟羽化盛期、孵化初期。9月8~11日,每只引虫灯日诱蛾量达到1000只左右,5代桑螟呈暴发趋势;但在9月12日后遭遇“海癸”台风,连续大风大雨,结果5代桑螟发生量比历年低,预期暴发并没有出现,少数地块出现较重危害,危害高峰期也比预测期推迟5 d左右。
桑螟属暴发潜力较大害虫,在自然环境中遇到适合环境条件,增加倍数可以达到30以上。据浙江湖州地区,1998年四代桑螟数量为每公顷21000条,处在较低水平,至5代时每hm2数量突然增到600000条以上,增长了30.7倍[23]。所以,常规“压前控后”的害虫防治技术,不适合桑螟防治策略。
2.3 农事操作
桑螟发生数量与农事操作有着密切关系。在春季,如果蚕桑生产布局合理,桑螟危害高峰出现之前,家蚕已经上蔟,通过夏伐,可以把大部桑螟带出桑园,桑螟虫口基数急剧下降。如果春蚕生产饲养延迟,在家蚕上蔟前,大批桑螟老熟化蛹,后代桑螟数量就大。在秋季,8~9月份是蚕桑生产中秋、晚秋饲养期,也是最适合桑螟发生期,如果养蚕生产布局合理,通过合理采摘桑叶,可以把大部分桑螟带出桑园,可以控制桑螟数量和灾害形成。如果桑螟危害高峰先于养蚕用叶高峰,桑螟的灾害就会形成[7]。如湖州地区在1997年和1998年调整秋蚕生产布局,把传统的早、中、晚3季秋蚕改为早中秋蚕和晚中秋蚕两期,发种时间是:早中秋8月20日和9月10日,与秋期四、五代桑螟发生期完全重合。结果连续两年5代桑螟大暴发,桑叶损失在15%以上,导致许多农户中晚秋蚕无法正常饲养。之后,对秋季养蚕布局又进行了调整,把中秋蚕发种时间调整到8月25日至9月1日之间,把晚秋蚕发种时间调整到9月20日至9月25日之间,结果从2000年至2012年五代桑螟一直没有出现严重危害。家蚕和桑螟都是鳞翅目昆虫,以桑叶为食,如果家蚕先食尽了桑叶,桑螟由于缺乏食料就不能顺利完成世代,所以,通过合理安排养蚕采叶、伐条等农事操作,可以调节桑螟发生数量,影响桑螟发生规律。
2.4 天敌昆虫
桑螟生存策略为r对策,由于众多天敌控制,使其数量受到压制;桑螟天敌有,卵期寄生蜂[28]:广赤眼蜂(Trichogramma evanescens Westwood),松毛虫赤眼蜂(TdendrolimiMatsumura)。由于桑螟卵很小,在大田调查中很少发现卵寄生蜂。
幼虫期寄生天敌有[28,29]:混腔室茧蜂(Aulacocentrum confusum He et van Aehterberg)桑螟绒茧峰(Apanteles helerusiae Wilkinson),甲腹茧峰(Chelonus sp.),菲岛愈腹茧(Phanerotoma philippinensys Ash-mead),红胸齿腿姬蜂(Pristomerus erythrothoracis Uchida),守子蜂(Cedria paradoxa W iakinson),菲岛长距茧蜂(Macrocentrus philippinensis Ashmead)。各寄生蜂都在桑螟1~4龄期开始寄生,有的能在桑螟幼虫暴食前致其死亡,有的则在桑螟老熟后致其死亡。
桑螟蛹期寄生蜂有3种[28~29]:广黑点瘤姬蜂(Xanthopimpla punctata Fabricius),体长10~12 mm,黄色。单寄生,跨期寄生于老熟幼虫至蛹,于桑螟蛹内化蛹、羽化。广大腿小蜂[Brachymeria lasus(Walker)]雌蜂体长5.2~6.5mm,雄蜂略小。单寄生,跨期寄生于老熟幼虫至蛹,在寄主蛹内化蛹、羽化。柄腹姬小蜂(Pediobirs sp。)体长1.5~2.3 mm,体淡绿色,有金属光泽。多寄生,从寄主蛹内羽化,1头桑螟蛹内可羽化出20~30头姬不蜂。蛹期寄生蜂田间寄生率较低,一般不超过1%。
重寄生蜂有3种[28]:中华横脊姬蜂(Stictopisthus chinensis Uchida)单寄生于桑螟绒茧蜂体内,由绒茧蜂体内羽化。次生大腿小蜂[Brachymeria secundaria(Ruschka)]。桑螟广肩小蜂(Eurytoma sp.)单寄生于桑螟绒茧蜂体内。
捕食性天敌有茶翅蝽,步行虫,黄蜂等。
桑螟虽然各时期都能被天敌寄生,但主要被寄生时期是幼虫期。据余虹、于香军等[28,30~31]调查,桑螟幼虫优势寄生蜂主要有2种,分别是混腔室茧蜂和桑螟绒茧蜂,并对这两种寄生蜂的生活习性和寄生规律进行了深入研究和详尽描述。
混腔室茧蜂:最高寄生率可达59%,平均寄生率为50.54%。其寄生方式是容性寄生,即被其寄生后的桑螟幼虫,在老熟前不表现任何症状,保持正常取食,直至老熟时其身体不显红色而呈乳白色,体缩短,用手挤压其腹部可见一内含物随之滚动,即寄生蜂的幼。不久,寄生蜂幼虫钻出寄主体外,并继续吸附于寄主幼虫体上取食,随着桑螟幼体逐渐干瘪,寄生蜂幼虫身体迅速膨大,然后叶丝结茧,此过程仅需1 d即可完成。
桑螟绒茧蜂:对各代桑螟的幼虫均能寄生,最高寄生率达到55.41%,平均寄生率为33.99%。绒茧蜂多寄生于桑螟1~2龄的幼虫,偶尔寄生3龄幼虫,而4、5龄幼虫已不适合绒茧蜂寄生。刺激桑螟绒茧蜂搜索寄主的利它素存在于寄主幼虫、为害叶、粪便之中,热稳定性好,能被甲醇、二氯甲炕、氯仿所抽提。被桑螟绒茧蜂寄生的桑螟低龄幼虫,体色由水绿色变成淡黄色,行动缓慢,发育延迟,至3龄末4龄初时,幼虫开始急躁好动,爬行迅速,可透视腹部内呈淡红色的桑螟绒茧蜂幼虫。不久,桑螟绒茧蜂幼虫从桑螟幼虫体内钻出,并附着在桑螟幼虫体外继续吸取寄主体液,直至桑螟幼虫干瘪死亡,桑螟绒茧蜂则在其附近的桑叶背面结茧化蛹。所以,被寄生的桑螟幼虫大部分死于4龄暴食前,对于控制当代桑螟的危害和次代桑螟发生具有重要意义。
据浙江湖州地区调查[7],桑螟天敌自然寄生率最高可达到70%以上。在桑螟数量最多的第4、第5代相对寄生率较低,翌年第1代桑螟时天敌寄生率开始回升,至第2代时寄生率达到高峰,第3代桑螟时天敌寄生率开始下降。即在桑园生态系统中,天敌与桑螟种群处在一个相对动态平衡之中。天敌随着桑螟种群的数量上升而上升,桑螟随着天敌的数量上升而下降,天敌下降而桑螟上升。天敌控制着桑螟,依赖于桑螟生存繁殖。天敌与桑螟种群的动态过程呈现出跟随现象。
2.5 化学农药
桑园在化学农药没有使用前,主要害虫是桑蟥、野蚕、桑毛虫,桑螟一直是次生害虫。化学农药引入到桑园中之后,考虑到家蚕饲养安全,使用的都是广普、高毒、短残留农药,桑蟥、野蚕逐渐被控制,目前危害很轻。但桑螟隐蔽性较强,折叶危害大,对桑蟥、野蚕高效的农药,对桑螟控制效果一般,对螟类高效的农药,由于其对家蚕残效期太长,在桑园中不能使用。并且,由于蚕农长期在养蚕间隙使用农药,使桑螟发生世代与家蚕饲养重合度很高,防治难度越来越大。桑螟是一类灾变能力极强害虫,在极短时间内能快速翻倍增长,在上一代桑螟虫源基数很低的情况下,遇到适合的环境条件,能迅速暴发成灾。所以,随着近年农药大量、不规范的使用,桑螟危害却在逐年加重,桑螟逐步成为桑园主要害虫。
3 防治技术
桑螟成灾特点:桑螟灾害形成不但在于桑螟数量,也和养蚕采叶有很大关系。桑螟食叶与家蚕食叶都有一个渐变过程,1~3龄食桑曲线平缓,4龄时开始转折,5龄达到高峰。所以,桑螟危害高峰先于养蚕用叶高峰,灾害就形成,如养蚕用叶高峰先于桑螟危害高峰,灾害损失就不会形成。
桑螟防治策略:桑螟防治,应从桑螟灾变合成条件入手,改变桑螟生存环境,辅之化学农药防治手段进行综合治理,可以收到事半功倍的效果[7]。
3.1 农业防治
通过合理规划农事操作,提高桑园科学栽培管理水平,可以收到很好防治效果。一是通过科学安排蚕种饲养布局,使养蚕用叶高峰先于桑螟危害高峰,在不使用化学农药的情况下,能起到很好调节桑螟种群的数量。比如,近年气温升高,春季桑螟出现时间提早,如果蚕种饲养沿袭传统习惯时间,就容易出现桑螟灾害性危害。所以,通过适当提早蚕种饲养时间,在桑螟危害高峰没有到来前,已通过养蚕大面积采叶和伐条,把大部分桑螟带出了桑园,防治效果远胜于化学农药;在非常适宜桑螟发生的中晚秋时期,可以预测出主危害代桑螟幼虫孵化和危害高峰期,通过调整中晚秋发种时间,使养蚕用叶高峰先于桑螟危害高峰,通过养蚕采叶,把桑螟带出桑园,桑螟发生数量虽没有减少,但不会造成桑叶实际损失,同时少用了一次蚕期农药,对确保秋蚕饲养安全有着积极意义,可以收到农药防治收不到的效果。二是科学安排全年蚕种饲养次数。近年在太湖流域蚕种养饲养次数越来越少,大部分农户夏蚕、早秋都不饲养。至中晚秋时,桑园非常茂盛,郁蔽度、湿度高,通风差,非常有利于桑螟繁殖。通过增养家蚕次数,采摘了下部桑叶,一则可带走下部桑螟,减少桑螟虫源基数,同时改善了桑园农事操作环境,有利于提高桑园化学农药防治质量。三是及时夏伐,春季余叶的桑园不能留养致秋期,这样减少了桑螟从春夏至秋的过渡性桥梁寄主,不仅减少了当代桑螟数量,也减少了下一代桑螟虫源基数。四是及时做好桑园的结束和解束工作。据调查,桑螟有79.74%在束草内越冬,2.26%在树穴内越冬,因此。在每年养蚕期结束后,及时在桑树上束草或桑园四周堆草,为桑螟老熟幼虫提供越冬场所。在翌年2~3月,幼虫开始活动前集中烧毁草束及堆草,可大幅度降低桑螟越冬虫口数,对防治桑螟有较好效果。五是建设健康型的桑园,单位面积内合理种植桑树株数,均衡施肥,桑园中沟渠通畅,清除桑园内的枯枝残叶,建立一个有利于降低桑螟繁殖系数、桑树健康生长的环境。
3.2 生物防治
桑螟天敌资源十分丰富,在桑园管理、采叶过程中可随时保护和利用天敌。如在采叶时,若发现被采桑叶上有桑螟绒茧蜂小白茧,应将其放在桑树上,让它羽化寄生,减轻桑螟的危害;在桑树落叶前或落叶时,收集桑叶上的小白茧,至翌年4月再将小白茧放回桑园,以抑制第1代桑螟的发生;应尽量少施用对该天敌有杀伤力的农药,以保护桑螟绒茧蜂。利用有利于天敌繁衍的耕作栽培措施,选择对天敌较安全的选择性农药,并合理减少施用化学农药,保护利用天敌昆虫来控制桑螟种群。
3.3 物理防治
桑螟成虫有着明显的趋光性,生产上可以利用频振式杀虫灯或黑光灯进行诱杀,从而有效降低成虫种群密度及后代发生数量。
3.4 化学防治
桑螟的防治指标为[32]:春夏季为24000~30000头/hm2,秋季为36000~60000头/hm2。防治桑螟的药剂种类较多,在桑树上登记、防治效果较好的种类及其使用浓度为:40%灭多威乳油2000倍液、8%残杀威可溶性粉剂1500倍液、40%毒死蜱乳油1500倍液、40%辛硫磷乳油1500倍液、60%敌·马合剂1000倍液、80%敌敌畏乳油800倍液。
当前,桑螟对桑园上常用的化学农药已产生较大抗药性。对灭多威抗性达到35.2倍;对双效磷抗性10.6倍,对敌敌畏抗性已达到很高水平,实验室检测的半致死浓度已高于大田防治的推荐浓度;对辛硫磷和毒死蜱的抗性在5倍左右,处于低水平抗性阶段。抗灭多威的桑螟品系对毒死蜱不存在交互抗性,所以,在对灭多威已产生很高抗性的地区,防治桑螟时可用毒死蜱与灭多威交替使用或镶嵌使用,延缓桑螟抗性发展[33]。
桑螟暴发能力强,常规“压前控后”防治技术对桑螟不是很适合,因为即使上代基础很低,只要环境条件适宜,就会暴发成灾。浙江湖州地区提出桑螟化学防治策略是“治好第1代,兼治2、3代,重点防治4、5代”。蚕种生产桑螟防治策略为:主要做好越冬代桑螟防治和清理工作。桑螟化学防治技术核心是:及时防治;在桑螟没有出现折叶危害的3龄前,及时进行化学防治,可以收到事半功倍效果[34]。
4 结束语
桑螟目前已成桑园主要害虫,桑螟灾害控制已从一项纯单纯的喷药防治技术演变为一项复杂的系统控制工程,其控害消灾的目标也应从喷药治虫提高到环境保护和蚕业可持续性的高度来认识,随着蚕桑生产结构调整和桑螟的演变,桑螟防治方法也在不断改进,对桑螟发生规律,特别是暴发规律和控制技术研究工作还任重道远,今后还需要多学科众领域的交
[1]黄尔田.实用桑树保护学[M].成都:四川科学技术出版社.1991:211~221.
[2]黄伟书,白丽涛.桑螟虫综合防治方法探讨[J].蚕学通迅,2008,28(3):32~34.
[3]钱祥明,洪志英,王卫明,等桑螟的生物特性研究[J].蚕业科学,1995,21(1):50~52.
[4]吴华,严余高,张春凤等.盐城市中部地区桑螟大暴发原因及其防治对策[J].江苏蚕业,2001,(2):24~25.
[5]王卫民明,方利民,朱引根等.桑螟暴发性特征指数分析与预测模型的研究[J].江苏蚕业,2007,(4):49~52.
[6]沈永泉,卢春松.桑螟暴发原因浅析及防治对策[J].蚕桑通报,1997.
[7]白锡川,杨海江,洪缨莉.桑螟发生规律及防治策略[J].昆虫知识,2002.,39(5):366~369.
[8]陈小青,朱方容.桑螟为害与发生规律的研究[J].广西蚕业,2006,43(3):29~33.
[9]莫现会,陈小青,林强,等.我所桑螟发生及为害调查初报[J].广西蚕业,2001,38(1):30~31.
[10]王文学,杨胜特,胡仕叶,等.遵义地区桑螟发生原因及综合防治技术[J].植物保护学.2012,11:137~139.
[11]方树友,滕淳茜,肖建京.淳安县桑螟加重发生的原因与防控对策[J].中国农技推广.2008,24(2):40~41.
[12]舒衷远,李政.桑螟虫发生为害规律及防治对策[J].四川蚕业.2000,(1):20~21.
[13]王文学,杨胜特,胡仕叶,等.遵义地区桑螟发生原因及综合防治技术[J].2012,(11):137~139.
[14]徐俊,卢鹏,余春华.修水县桑树主要病虫害发生特点及防治方法[J].植物保护与推广.2001,21(7):20~21.
[15]羿红.桑螟的发生与防治研究[J].蚕业科学,2000,26(3):182~183.
[16]白锡川,杨海江,陆鸿英,等.光周期对浙北地区桑螟滞育的影响[J].蚕业科学.2002,28(4):329~332.
[17]王敦崇,张岩.越冬代桑螟羽化规律调查与分析[J].江苏蚕业.2007,(2):20~21.
[18]王建新.桑树害虫的系统预测及其方法[M].杭州:杭州大学出版社.1992.
[19]谢立群,毛建萍,浦冠勤,等.桑螟实验种群生命的研究[J].蚕业科学.2003,29(3):222~225.
[20]何春华,金一林.桑螟饲养方法及发生规律初探[J].蚕桑通报.1990,20(2):20~22.
[21]东永杰,孙绪艮,张卫光,等.桑螟越冬幼虫体内水分、脂肪、甘油的变化与抗寒性的关系[J].蚕业科学.2005,31(1):22~25.
[22]东永杰,孙绪艮,张卫光,等.桑螟越冬幼虫体内蛋白质、氨基酸、碳水化合物的变化与抗寒性的关系[J].蚕业科学.2005,31(2):111~116.
[23]白锡川,杨海江,陆鸿英,等.1998年桑螟消长动态分析及防治[J].蚕桑通报.1999,30(2):20~23.
[24]刘康成,杨志华,习平根,等.桑螟发育起点温度和有效积温的研究[J].昆虫知识.1996,33(2):85~87.
[25]张建强.桑螟实验种群数量动态研究[J].蚕业科学. 1990,16(2):108~110.
[26]谢立群,王卫明,浦冠勤.桑螟实验种群数量变动的模拟[J].蚕业科学.2002,28(3):261~264.
[27]毛美红,白锡川.气象要素的时间分布对五代桑螟数量影响分析[J].昆虫知识.2009,46(1):56~60.
[28]余虹,周勤.浙江省桑螟寄生蜂调查研究[J].蚕业科学. 2003,29(4):330~334.
[29]于军香,毛建萍,谢立群,等.桑螟寄生性天敌昆虫的研究[J].蚕业科学.2002,28(4):273~276.
[30]余虹,周勤.宋毓.桑螟绒茧蜂的生物学特性[J].浙江大学学报.2004,30(5):557~560.
[31]于香军,毛建萍,谢立群,等.桑螟绒茧蜂的生物学特性[J].昆虫知识.2005,42(2):199~201.
[32]浦冠勤,毛建萍,于军香,等.桑螟食叶量测定及其防治指标的研究[J].蚕业科学.2004,30(2):207~210.
[33]白锡川,吕美坤,张德明.桑螟对桑园常用农药抗性调查[J].植物保护.2005,31(6):81~83.
[34]白锡川,杨海江,陆鸿英.桑螟化学防治策略[J].蚕桑通报.2001,32(2):22~24.
Research Progress on Outbreak Regu larity of Mulberry Moth and Control Techniques
ZHAO Xu-hua1,BAIXi-chuan2,SH IGuo-fang1
(1.Huzhou Academy of Agricultural Sciences,Huzhou 313000 Zhejiang,China; 2.Wuxing Agricultural Technology Extension and Service Center,Huzhou 313000 Zhejiang,China)
The mulberry moth has gradually become the primary pest in mulberry fields because of its wide spread, strong fecundity capacity,terrible diffusion ability.This articlemainly summarized the research progress of outbreak regularity,life habit and control technique ofmulberry moth.
mulberrymoth;outbreak regularity;control technique
S888.7
A
0258-4069[2013]03-005-06
赵叙华(1959-),男,农艺师。主要从事蚕桑技术推广工作。E-mail:zhaoxuhua1959@163.com
