猪对“非必需氨基酸”的营养需要
2013-02-20伍国耀武振龙戴兆来王薇薇刘王军军印遇龙
■ 伍国耀 武振龙 戴兆来 杨 鹰 王薇薇刘 闯 王 斌 王军军 印遇龙
(1.德克萨斯州农工大学动物科学系,德克萨斯州,美国 77843;2.中国农业大学动物营养学国家重点实验室,北京 100193;3.中国科学院动物营养与健康实验室和农业生态重点实验室亚热带农业生态研究所,湖南长沙 410125)
提供足够的日粮AA是动物健康、生长、发育和存活必不可少的。基于猪的生长或者氮平衡,传统上把AA分为必需氨基酸(EAA)和非必需氨基酸(NEAA)(见表1)。EAA是其碳骨架在动物细胞内不能合成或合成速度不能满足最佳生长、发育和健康需要,必须由饲粮提供来满足动物需要的氨基酸。相反,NEAA是在动物体内能够充足合成且可以满足动物维持、生长、发育和健康需要的氨基酸,因此,不需要通过日粮提供。迄今为止,没有令人信服的证据证明在猪营养中未被纳入EAA类的所有氨基酸能够在其体内充足的合成。显然地,EAA和NEAA术语的定义是存在问题的。然而,在“非必需营养”理念的指导下,养猪生产者在日粮配方中长期忽视NEAA。本文的主要目的是引起读者们注意NEAA的定义在理论和实践上的局限性。

表1 猪营养中将AA分为EAA和NEAA的传统分类
1 NEAA营养的生理作用
AA在动物生理上有多个角色(图1)。首先,AA调节基因表达(翻译基因编码为核糖核酸和蛋白质),核糖核酸RNA,包含信使、转录和核糖体RNA。这是一种高度特异性过程:一种基因在其特异性调节因子作用下被打开或关闭。除了作为蛋白质合成的底物,AA可影响以下步骤的一个或多个:染色质修饰(DNA复合物和覆盖蛋白质,例如组蛋白)、转录(从DNA合成mRNA)、转录后修饰、RNA转运、mRNA降解、翻译(从mRNA合成蛋白质/多肽)和翻译后修饰。研究最多的4个NEAA是精氨酸、谷氨酰胺、甘氨酸和脯氨酸。例如,精氨酸激活雷帕霉素靶蛋白 (MTOR)信号途径,刺激仔猪肌肉蛋白的合成。同样,我们关于在早期断奶仔猪日粮中补充或不补充谷氨酰胺的基因芯片研究结果表明:早期断奶导致与氧化应激和免疫激活相关基因的表达增加(52%~346%),但是与营养素代谢和肠道细胞增殖有关的基因表达却降低(35%~77%)。日粮中补充谷氨酰胺增加了(120%~124%)细胞生长和去除氧化剂所必需的相关基因在肠道的表达,降低了(34%~75%)促进氧化应激和免疫激活的基因的表达。另外,谷氨酰胺加强了骨骼肌和小肠中MTOR信号和蛋白质的合成。这些发现揭示了断奶会引起某些基因表达的变化,并在分子机制上解释了日粮中补充谷氨酰胺有助于改善幼龄哺乳动物营养状况这一现象。

图1 NEAA在猪上的生理功能
第二,NEAA调节一氧化氮(NO)、一氧化碳(CO)和硫化氢(H2S)的合成,这些物质通过cGMP和cAMP的产生参与在细胞中的信号气体途径,从而加强血液流量、营养运输、蛋白质沉积和免疫功能。特别是,甘氨酸是血红素的前体,其氧化产生CO,而胱氨酸是H2S产生的底物。精氨酸通过NO合成酶(NOS)三个异构体中的一个合成NO,三个异构体为:神经元NOS(nNOS;也被称为 NOS1),可诱导NOS(iNOS;也被称为 NOS2)和内皮 NOS(eNOS;也被称为NOS3)。许多NEAA被报道通过细胞依赖性方式调节NO、CO和H2S的产生。例如,精氨酸、瓜氨酸、谷氨酸、甘氨酸、牛磺酸和γ-氨基丁酸通过在内皮细胞或者脑中的组成型NO合成酶增加NO的合成,但是谷氨酰胺却通过组成型NO合成酶和诱导型NO合成酶两种方式调制NO的产生。此外,精氨酸、谷氨酰胺、谷氨酸、丙氨酸、牛磺酸和甘氨酸通过内皮细胞核非血管组织的血红素加氧酶促进CO的合成,但是N-乙酰基-半胱氨酸减少受损的脑和血管平滑肌细胞中CO的形成。因此,NO、CO和H2S产生不足将会损害整个机体胰岛素的敏感性和降低营养素用于蛋白质在组织中沉积的效率。
第三,NEAA(例如精氨酸、谷氨酸、谷氨酰胺和脯氨酸)参与细胞信号途径包括MTOR、依赖cAMP的蛋白激酶、依赖cGMP的蛋白激酶、G-蛋白偶联受体、AMP激活蛋白激酶(AMPK)、有丝分裂原活化蛋白激酶(MAPK)。这是一个复杂的代谢调控网络,但是他们都受蛋白磷酸化和蛋白去磷酸化机制调节。这些途径还涉及在细胞质和细胞核中的各种蛋白质的活化,从而调节细胞活动的生理过程,包括:基因表达;营养代谢;细胞增殖、分化和迁移;有丝分裂和细胞存活;细胞循环周期;细胞存活和凋亡;炎症反应。
第四,NEAA是合成许多具有重要功能的含氮物质的底物。表2列出了这些生物活性分子中的一部分,包括肌酸,谷胱甘肽、神经递质和低分子量的激素。这些物质是猪生长、泌乳、繁殖、健康和存活所必需的。如:①营养素的吸收和代谢[例如:养分运输、蛋白质周转、脂肪合成和氧化、葡萄糖合成和氧化、氨基酸合成和氧化、氨解毒作用(合成尿素和尿酸)、食物利用效率];②调节内皮细胞的功能、血流量、淋巴循环以及免疫功能和健康(例如:T-细胞增殖和B-细胞成熟、B-细胞产生抗体、杀灭病原体、肥胖、糖尿病和代谢综合症);③精子的产生、雄性的繁殖力、排卵,卵巢甾体激素合成,胚胎植入和存活、胎盘血管生成和生长,胎儿的生长发育和泌乳能力;④酸碱平衡、神经传导,细胞外和细胞内的渗透压,抗氧化防御和体内平衡;⑤产后的存活、生长和发育,以及组织再生和重塑。
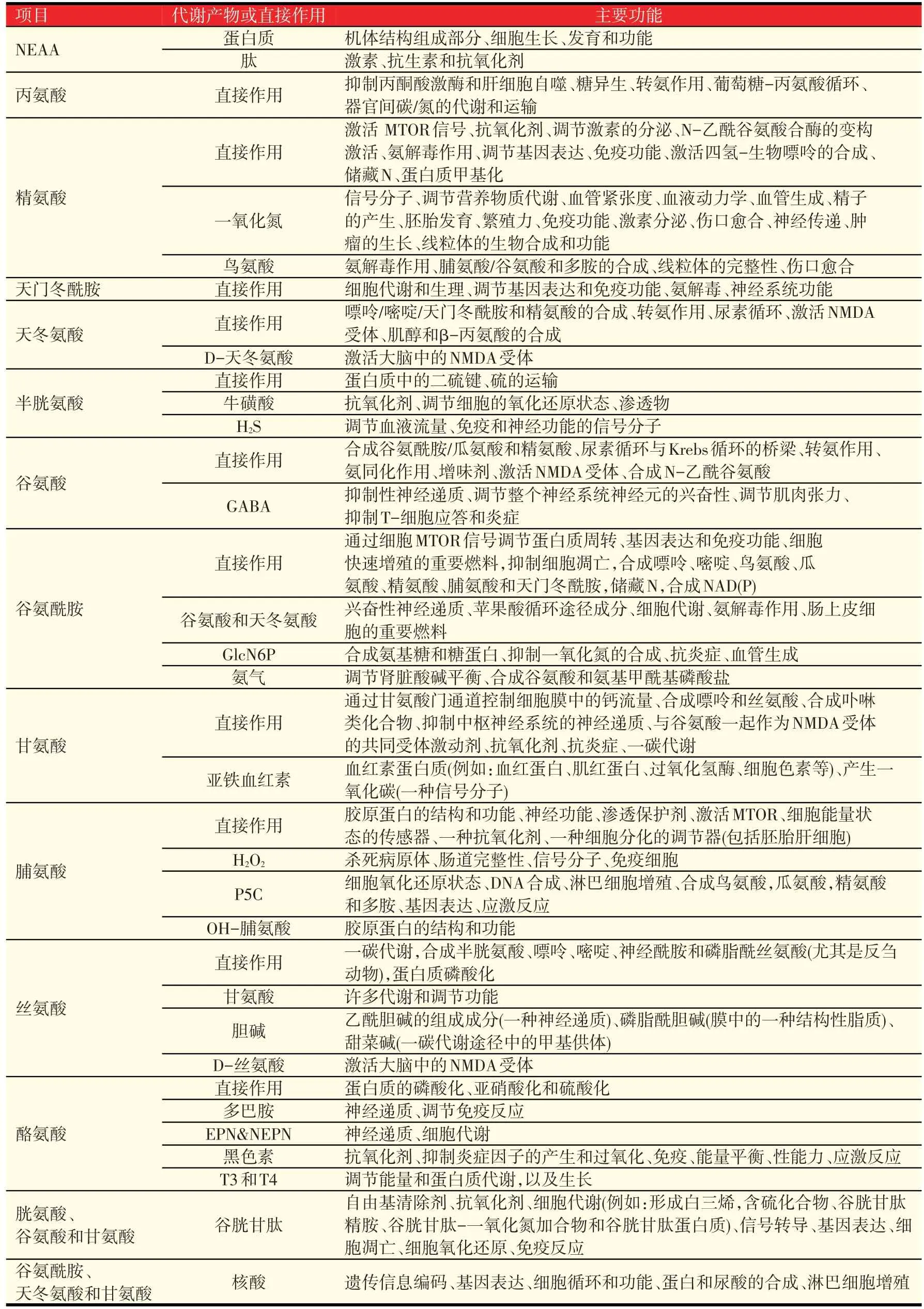
表2 NEAA在营养和代谢上的主要代谢产物和功能
2 日粮NEAA的需要
日粮NEAA的需要应根据所有AA在维持、组织蛋白合成、生理上重要的非蛋白代谢产物的产生和它们的调节功能的代谢需要等方面来确定。因此,日粮NEAA的需要可能随着营养、生理、病理和环境因素变化而不同,应该包含在“理想蛋白质”理论里(图2)。Deng等报道,虽然幼龄仔猪日粮中含有相同数量的EAA,日粮中NEAA的不足会使猪的组织蛋白合成和生长性能受到限制(表3)。尽管认可NEAA在动物生长、泌乳、繁殖和健康方面具有重要的作用,但是最新版本的美国国家研究委员会(NRC)猪营养需要并没有在猪任何生长阶段的日粮需要中推荐NEAA。NRC表明,猪具有足够合成NEAA的能力。然而,最近的研究证据与这个理念是不一致的。此外,动物机体NEAA的合成非常依赖昂贵蛋白质原料中EAA的含量,在生产上为了降低成本而给猪饲喂普通的日粮,其EAA并不一定能用来合成充足的NEAA。这就说明根据AA代谢领域新发展的最新的知识推荐AA需要量而保证日粮中NEAA的需要是非常重要的。
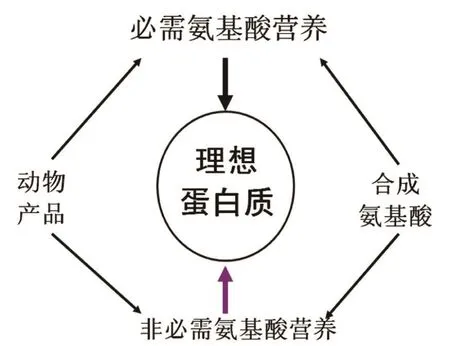
图2 推荐猪日粮NEAA的需要来修正“理想蛋白质”概念
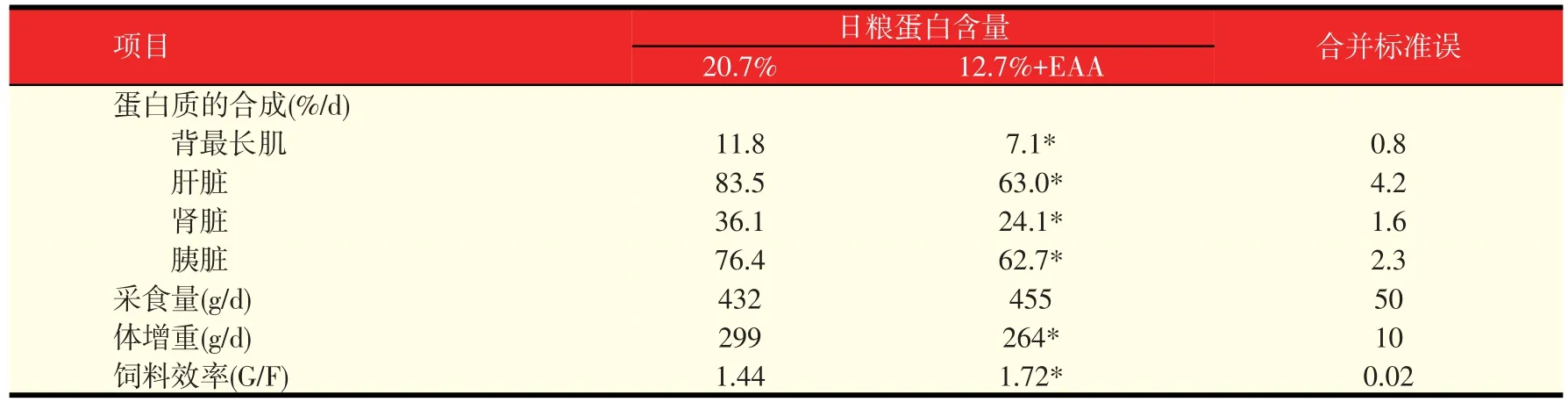
表3 低蛋白日粮缺乏NEAA限制了仔猪(25~39 d)组织蛋白质的合成和生长
事实上,一些AA在体内的合成会消耗大量的能量来体现它们在生理上的重要性。因此,这些AA在体内的从头合成途径已经发生演变或者高度保守。此外,所有的NEAA经过代谢转换后而具有重要的生理功能。例如,出生后的仔猪的血浆中通常含有高浓度的(~1 mM)甘氨酸在刺激仔猪快速生长方面具有重要的作用,早期妊娠猪尿囊液中丰富的精氨酸(达到6 mM)促进胎盘生长和胎儿发育。因此,有令人信服的证据表明,日粮中NEAA不足会影响猪的免疫和生产性能。例如,最近的研究表明:①日粮必须含有足够量的精氨酸和谷氨酰胺来维持猪在胎儿期、新生儿期和断奶后的生长和抗病能力;②日粮补充脯氨酸或者谷氨酸可提高早期断奶仔猪的生长性能和饲料效率。根据奶蛋白基础日粮中苯丙氨酸在体内氧化的数据,Ball等建议将脯氨酸作为幼龄仔猪的EAA。
猪日粮NEAA的需要量应根据生产者们期望的猪的生长和生产性能水平来推荐。根据已发表的研究,表4列出了我们用传统的玉米-豆粕型基础日粮饲喂怀孕、泌乳母猪以及断奶仔猪和生长-育肥猪时的精氨酸、谷氨酰胺、谷氨酸和脯氨酸的推荐量。当使用新原料时,应确定其在不同发育阶段猪上的日粮AA的真消化率。可以通过添加合成AA或者含有丰富的EAA和NEAA的动物源产品来补充NEAA,从而满足动物的需要。表5列出了不同原料中的各种AA组成成分,便于在配制日粮配方时满足所有AA包括EAA和NEAA的需要。
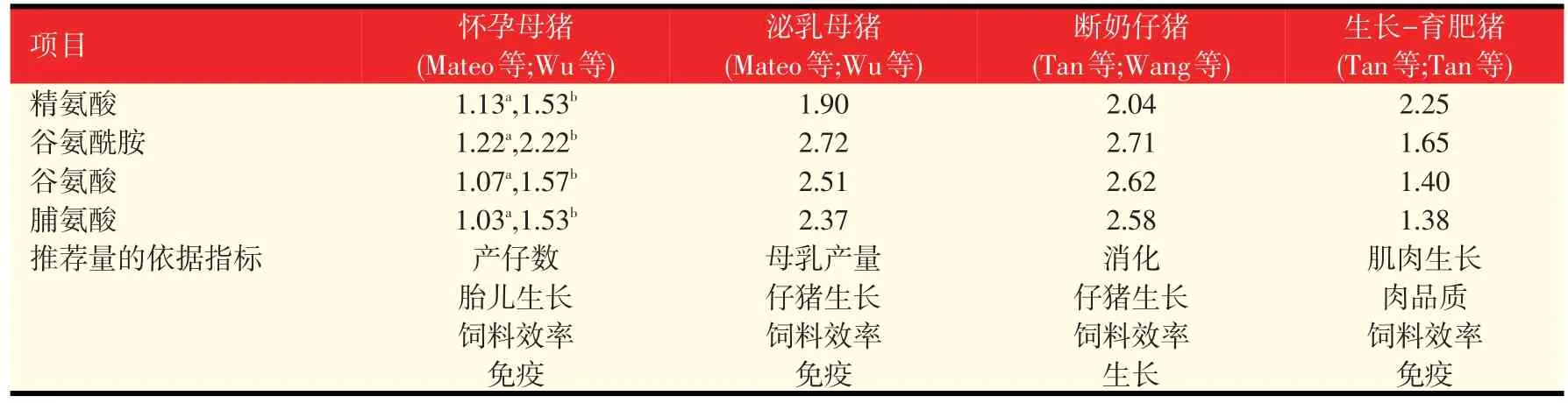
表4 猪日粮中NEAA需要的推荐量(%)(以干物质为基础,玉米-豆粕型基础日粮)
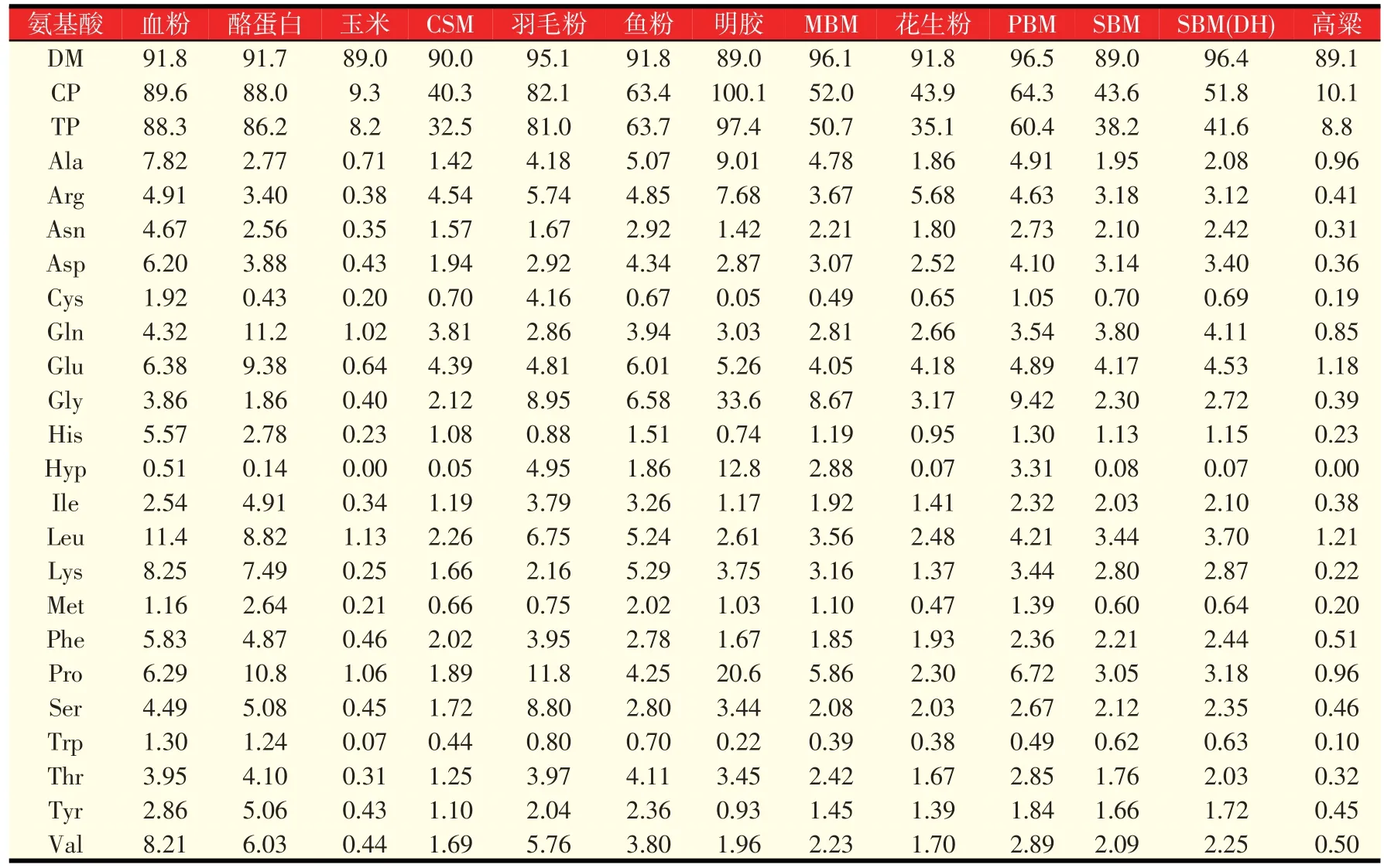
表5 饲料原料中总AA的组成(%,以干物质为基础)
3 结论
将AA分为EAA和NEAA的传统分类法在猪蛋白质营养上存在着概念上的限制。遗憾的是最新版的NRC仍然没有提供猪NEAA的日粮需要推荐量。然而,新的证据表明被传统分类为NEAA的氨基酸,尤其是谷氨酰胺、谷氨酸、脯氨酸和精氨酸在转录和翻译水平调控基因表达中发挥了重要的作用。人们越来越认识到NEAA通过MTOR,AMPK、MAPK和气体(NO、CO和H2S)参与细胞信号途径。这些调控网络的完全整合对细胞增殖、分化、代谢、平衡、生存和功能具有重大的影响。因此,经典的“理想蛋白质”概念应该包含EAA和NEAA,从而在全球畜牧业上提高猪利用饲料的效率及其生产性能。
(参考文献42篇,刊略,需者可函索)
