局部表象产生中行为性别差异的神经基础*
2013-02-03赵庆柏张小菲隋丹妮周治金陈其才周宗奎
赵庆柏 张小菲 隋丹妮 周治金 陈其才 周宗奎
(1青少年网络心理与行为教育部重点实验(华中师范大学),武汉 430079)(2人的发展与心理健康湖北省重点实验室,华中师范大学心理学院,武汉 430079)(3沈阳大学外国语学院,沈阳 110044) (4华中师范大学生命科学学院,武汉 430079)
1 引言
表象作为重要且基本的认知能力之一,是由多个子系统构成的复杂的心理过程,包括表象的产生、扫描、保持、旋转等过程,其中最基本的是表象的产生。表象的产生是指通过重新激活存贮在长时记忆中的信息,而产生瞬间的内在视觉表征的认知过程(Kosslyn,1994)。研究者多采用文字诱发范式来研究视觉表象产生,即给被试呈现线索词,让其产生相应的心理图像(De Beni,Pazzaglia,&Gardini,2007)。
视觉表象与视觉感知机能等价(Kosslyn,1994),而整体优先性是视觉感知的一个重要特征(Navon,1977)。隋丹妮等人(2007)采用整体-局部范式对视觉表象产生是否存在整体优先性进行了研究。首先让被试习得20张动物图片,然后要求被试根据提供的词汇线索产生相应心理图像。词汇线索分为两种类型,分别指向先前习得图片的整体和局部(如大象—— 象鼻)。结果发现,整体优先性同样存在于表象产生中。此外,研究还发现在表象产生中存在着性别差异,即女性产生图像局部特征的能力要优于男性,而在产生整体轮廓特征时无显著性别差异。一般地,男性在视觉空间定位、整体-局部视知觉加工以及心理旋转等空间视觉能力上表现出优势(Basso& Lowery,2004;Roalf,Lowery,&Turetsky,2006;Voyer&Hou,2006);而女性却具有更好的言语、精细动作技能以及情景记忆等能力(Basso,Harrington,Matson,&Lowery,2000;Herlitz,Nilsson,&Backman,1997;Peters&Servos,1989)。由定义可知,记忆在表象产生中发挥了重要作用。最近一项研究(Harness,Jacot,Scherf,White,&Warnick,2008)表明,女性较男性具有更好的视觉(图片)工作记忆能力。这可能是导致女性在局部表象产生中反应更快的原因之一。
然而,这些结果多是基于行为反应时的。那么,这种行为反应上性别差异的神经基础是什么呢?一般地,反应时被认为是代表大脑信息加工速度的一个指标(Jensen,2006)。换句话说,任务状态下大脑信息加工速度决定了反应时的长短。根据大脑功能的两种基本组织模式—— 功能特化与功能整合(Tononi,Sporns,&Edelman,1994),大脑信息加工可视为两个层面:与不同子功能相对应的局部脑区的激活以及这些脑区间的相互作用。
在功能特化层面,Han等人(2002)采用功能核磁共振成像技术研究整体-局部视觉加工,发现当注意加工呈现在视野中央的复合刺激的整体性质时,右侧枕叶的活动加强;而当注意指向加工局部性质时,左侧枕叶的活动加强。基于事件相关电位的研究(Han,He,&Woods,2000)也显示,在加工整体性质的条件下,会在右半球引起较大幅度的与靶目标加工相关的差异波,但在加工局部条件时差异波主要分布在左半球。这些结果表明,左半球为加工局部信息的优势半球。此外,性别间表现出不同的半球偏侧化,其中女性为左半球优势(Kaiser,Kuenzli,Zappatore,&Nitsch,2007;Tomasi&Volkow,2012)。据此,隋丹妮等人(2007)采用半球不对称性来解释局部表象产生中的行为性别差异,认为女性是典型的左半球加工模式并且左半球被认为是加工局部信息的优势半球,因此在局部表象产生中女性反应更快。然而,此解释的依据主要是源自视觉感知,而非视觉表象,因此只是间接的推测,目前尚无直接的证明。
在功能整合层面,网络分析,特别是小世界网络(Watts&Strogatz,1998)被广泛用于评估大脑系统的信息加工。大量研究(Bullmore&Sporns,2009)发现,结构及功能脑网络均表现出小世界拓扑,即较大的簇系数和较短的平均路径长度。簇系数代表网络的局部效率,描述的是该节点被移除后其邻居间的信息流通能力(Latora&Marchiori,2001)。此外,大的簇系数能够提高空间分离的神经团之间的功能重叠(Sporns&Zwi,2004)。而平均路径长度则代表了信息的全局整合。较短的平均路径长度意味着较少的信息加工步骤,在信息传递中较少的时间消耗(Kaiser&Hilgetag,2006)。因此,脑网络的小世界拓扑为并行信息加工提供了较高的全局和局部效率(Achard&Bullmore,2007)。小世界网络不仅能为大脑内信息加工提供理论上的模型,同时能够为具体的研究提供量化的参数。
本研究采用事件相关电位(ERP)技术和小世界网络分析方法,对局部表象产生中性别差异的神经基础进行探索,并对行为表现与大脑信息加工模式之间的关系进行深入的分析讨论。
2 方法
2.1 被试
28名大学生被试参加了实验(男女各14人,女性年龄22.8±0.6岁,男性年龄23.3±0.8岁),均为右利手,身体健康,视力或者矫正视力正常。实验前均填写知情同意书。本次实验中所有被试的数据均有效。
2.2 设计
采用2(表象类型:整体表象和局部表象)×2(性别:男性和女性)混合实验设计,其中,表象类型为组内变量,性别为组间变量。实验以反应时、ERP波幅和功能脑网络的小世界参数(簇系数和平均路径长度)为因变量。
2.3 材料
实验材料是从200幅日常生活用品或电器中选用的60幅灰度矢量图片(Snodgrass and Vanderwart’s object databank),如电话、台灯等,图片大小8.5 cm×8.5 cm,每幅图片选定3个局部特征(文字描述,如灯罩、底座、支架等)。200位随机选取的大学生(全未参与后边的正式实验),仔细观察所给200幅图片,然后采用问卷调查的方式,对图片命名,对其熟悉度进行打分(0~4);并选出每幅图片中最突显的局部特征。200幅图片熟悉度平均得分为3.24,选取熟悉度大于平均值且单一局部特征指出率超过70%的60幅图片作为正式实验的材料,其中指出率最高的特征作为该图片的局部特征。实验图片统一为黑色背景,能充分展示物体的整体和局部特征。被试与屏幕的距离为60~70 cm,水平和垂直视角分别为8.0º~9.0º和6.0º~7.0º。
2.4 程序
被试坐在光线适度且无任何噪声的实验室内完成整个实验程序。实验开始,被试阅读屏幕上的指导语,并要求对其进行解释。在此过程中,任何误解均会被更正。呈现刺激材料的显示器背景为黑色,刺激位于方框中央。为了避免过多的脑电伪迹产生,要求被试在实验中始终注视显示器屏幕中央,尽量减少眼动。正式实验前有6个练习trial,以使被试熟悉实验任务。
正式实验流程如图1所示,每个trial开始,屏幕中心出现一个注视点,持续时间随机为1000 ms至2200 ms间的任一时间,以避免被试的期待效应。接着呈现图片2000 ms,之后是1000 ms的空屏,以消除视觉感觉记忆的影响。然后呈现与刚才图片整体或局部信息相关的线索文字(例如,整体线索:台灯;局部线索:灯罩)诱导被试在限定时间4000 ms内产生相应的表象(整体或局部)。一旦被试完成表象便按键,其行为反应时被同时记录。整个实验包含120个刺激(60幅图片,每幅出现两次,分别对应表示整体和局部信息的线索文字),所有刺激随机排列。

图1 实验程序流程图
2.5 ERP数据记录
使用德国Brain Products公司的脑电记录与分析系统,按国际10-20系统扩展的64导电极帽记录脑电活动,以FCz为参考电极,以位于Fpz上方AF3和AF4之间的电极接地,双眼外侧安置电极记录水平眼电(HEOG),左眼上下安置电极记录垂直眼电(VEOG)。每个电极处的头皮电阻保持在5 kΩ以下。滤波带通为0.05-100 Hz,采样频率为500 Hz/导。
完成连续记录EEG后离线处理数据,转换为双耳乳突平均参考,校正眼动及其他伪迹。根据被试的反应结果,只对表象成功产生(4000 ms内做出按键)的EEG进行叠加。波幅大于±80 μV 者被视为伪迹自动剔除。分析时程为线索文字出现前200 ms至其出现后800 ms,基线为前200 ms。
2.6 网络构建及分析
网络由节点和边构成。选取以下60个电极作为构建全脑功能网络的节点:Fp1,Fp2,AF7,AF3,AF4,AF8,F7,F5,F3,F1,Fz,F2,F4,F6,F8,FT7,FC5,FC3,FC1,FC2,FC4,FC6,FT8,T7,C5,C3,C1,Cz,C2,C4,C6,T8,TP9,TP7,CP5,CP3,CP1,CPz,CP2,CP4,CP6,TP8,TP10,P7,P5,P3,P1,Pz,P,P4,P6,P8,PO7,PO3,Poz,PO4,PO8,O1,Oz和O2。以电极之间的功能连通性作为网络的边。以前的研究(Nittono,Suehiro,&Hori,2002;Wu,Mai,Chan,Zheng,&Luo,2006)发现,表象效应发生在刺激呈现后的300-650ms,因此选取此时间段的ERP时间序列用于功能连通分析。
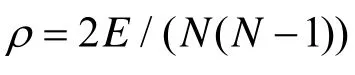
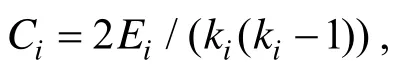
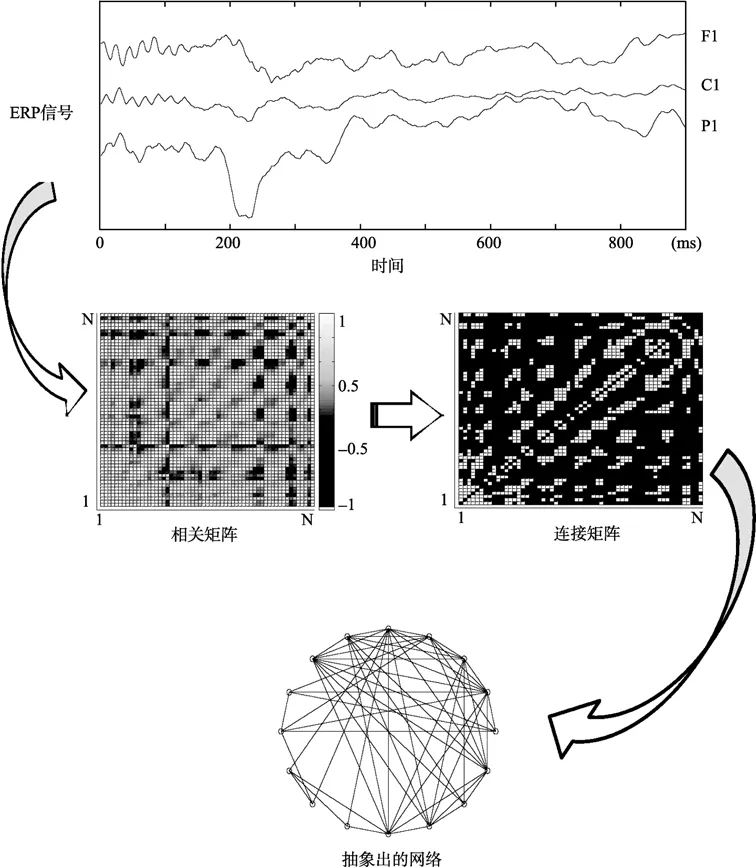
图2 功能脑网络构建示意图。
3 结果
局部表象产生中反应时的性别差异被再次验证。反应时分析显示,表象类型(整体、局部)主效应显著,F
(1,26)=18.21,p
<0.001;整体表象产生(M
=950.44,SD
±186.29)快于局部表象产生(M
=1053.64,SD
±203.90)。此外,表象类型与性别的交互作用显著,F
(1,26)=14.45,p
<0.001。简单效应分析发现,在整体表象产生中男女没有显著差异(p
=0.87),而在局部表象产生中女性显著快于男性(p
<0.05)(见图3)。这与先前的报道(隋丹妮 等,2007)是一致的。
图3 整体-局部表象产生中的反应时
ERP结果显示,在整体和局部表象产生中均存在明显的P1、N2以及一个300 ms以后的正成分(见图4)。分别对P1、N2和P300-650的平均波幅进行2(表象类型:整体、局部)×2(性别:男、女)×60(电极位置:见网络构建部分)的重复测量的方差分析,结果显示:(1)在P1平均波幅上,除电极位置主效应显著[F
(59,1534)=52.64,p
<0.001]外,其他主效应及交互作用均不显著;(2)在N2平均波幅上,除电极位置主效应显著[F
(59,1534)=27.82,p
<0.001],其他主效应及交互作用均不显著;(3)在P300-650平均波幅上,除电极位置主效应显著[F
(59,1534)=31.46,p
<0.001],任务类型与性别的交互作用显著[F
(1,26)=4.83,p
<0.05]外,其他主效应及交互作用均不显著。简单效应分析显示,局部表象产生中P300-650的平均波幅女性小于男性(p
=0.08),整体表象中无显著性别差异(p
=0.57)。对ERP数据进行对空间连通模式的分析,比较男女两组功能脑网络的簇系数及平均路径长度。如图5所示,在整体表象产生中,功能网络的簇系数和平均路径长度均无显著差异;在局部表象产生中,功能脑网络的簇系数无显著的性别差异,而平均路径长度女性较男性显著更短。
行为结果与功能脑网络的平均路径长度表现出一致性。进一步对二者相关性进行分析,结果显示,在局部表象产生中,反应时与平均路径长度成正相关(连边密度0.15~0.18范围内边缘显著,见表1),而其与簇系数则无显著的相关性。

图4 整体和局部表象产生的ERP波形图

图5 整体和局部表象产生中,相同连边密度ρ下男性与女性功能脑网络参数(簇系数C和平均路径长度APL)的比较

表1 局部表象产生中反应时与脑网络簇系数和平均路径长度的相关系数r及p值
4 讨论
表象产生中的行为性别差异得到了再次验证,即局部表象产生中女性显著快于男性,而整体表象产生中则无性别差异。本研究从功能特化和功能整合两个层面对这一行为差异的神经基础进行了探讨。
ERP结果显示,整体和局部表象产生均诱发了一个300~650 ms的正成分,与听觉表象产生的时间进程是相似的(Wu et al.,2006)。我们认为这可能是一个P3b成分,它反映的是认知资源的分配(Kok,2001)。任务分配的认知资源越多,其波幅越大。在局部表象产生中,女性的P3b波幅小于男性,且反应时短于男性,说明女性只需要较少的认知资源便可更好的产生局部表象。这与神经效率假说(Micheloyannis et al.,2006b)是一致的,即程度较低但相对集中的皮层激活反映了较高的神经效率。据此可推测,女性在局部表象产生中大脑信息加工效率更高。
功能脑网络分析的结果显示,簇系数在局部表象产生中没有表现出性别差异,与行为结果并不一致。一般而言,簇系数是局部连接的一个表征(Ferri et al.,2007)。本实验的结果意味着,大脑信息加工的局部连接模式并不是影响局部表象产生行为表现的重要因素。然而,平均路径长度则表现出与行为反应一致的特征,即女性在局部表象产生中具有更短的平均路径长度。在解剖脑网络中,短的平均路径长度赋予系统快速反应的能力(Lago-Fernandez,Huerta,Corbacho,&Siguenza,2000);而在功能脑网络中,它代表的是较强的全局信息加工整合能力。尽管功能连通性并不是直接描述真实的信息传递,但它能间接地反映脑区间的信息加工。研究结果显示平均路径长度与反应时间表现为正相关,正是对这一说法的支持。虽然相关性只是边缘显著,但这可能与被试数量有关。如果增加被试,相关应该可以达到显著性水平(p
<0.05)。在以往的报道中,小世界方法被广泛应用于神经精神疾病的临床研究中。研究者(He,Chen,&Evans,2008;Liu et al.,2008;Micheloyannis et al.,2006a;Rubinov et al.,2009;Supekar,Menon,Rubin,Musen,&Greicius,2008)发现,基于功能核磁共振成像或脑电的功能脑网络的小世界特性在精神分裂症和老年痴呆症中均表现出减弱。这预示着,小世界特性可能为临床提供了一种有用的诊断依据。此外,小世界方法也被应用于研究正常被试的认知加工(De Vico Fallani et al.,2008;Wang,Li,Metzak,He,&Woodward,2010)。在本研究中,小世界方法被成功用于量化局部表象产生中脑功能连通性的性别差异。并且研究发现了脑功能加工模式与行为反应间的关系,即短的平均路径长度支持了更快地行为反应。
由于电极定位的松散性和事件相关电位的多源性,研究只是为脑功能连通模式是行为反应的神经基础这一观点提供了间接支持,而不是直接的证据。今后的研究中,推荐采用高空间分辨率的功能核磁共振成像技术。此外,今后需要对各种不同认知任务进行研究,来验证反应时与功能连通模式这种关系的稳定性。
5 结论
实验从行为和神经生理两个层面对局部表象产生中的性别差异进行了深入探讨。在反应时上,女性具有更快的局部表象产生能力;在ERP成分上,对于局部表象产生诱发的P300-650女性具有更小的平均波幅;在脑功能网络参数上,局部表象产生的脑网络中女性具有更短的平均路径长度,且反应时与平均路径长度表现出正相关(边缘显著)。由此可推测,优化的大脑信息加工模式是快速行为反应的神经基础。
Achard,S.,&Bullmore,E.(2007).Efficiency and cost of economical brain functional networks.PLoS Computional
Biology,3
(2),e17.Basso,M.R.,Harrington,K.,Matson,M.,&Lowery,N.(2000).Sex differences on the WMS-III: Findings concerning verbal paired associates and faces.Clinical Neuropsychology,14
(2),231–235.Basso,M.R.,&Lowery,N.(2004).Global-local visual biases correspond with visual-spatialorientation.Journalof Clinical and Experimental Neuropsychology,26
(1),24–30.Bullmore,E.,&Sporns,O.(2009).Complex brain networks:Graph theoretical analysis of structural and functional systems.Nature Reviews Neuroscience,10
(3),186–198.DeBeni,R.,Pazzaglia,F.,& Gardini,S.(2007).The generation and maintenanceofvisualmentalimages:Evidence from image type and aging.Brain Cogn,63
(3),271–278.De Vico Fallani,F.,Astolfi,L.,Cincotti,F.,Mattia,D.,Marciani,M.G.,Gao,S.K.,...Babiloni,F.(2008).Structure ofthe corticalnetworks during successful memory encoding in TV commercials.Clinical Neurophysiology,119
(10),2231–2237.Ferri,R.,Rundo,F.,Bruni,O.,Terzano,M.G.,&Stam,C.J.(2007).Small-world network organization of functional connectivity of EEG slow-wave activity during sleep.Clinical Neurophysiology,118
(2),449–456.Ferri,R.,Rundo,F.,Bruni,O.,Terzano,M.G.,&Stam,C.J.(2008).The functional connectivity of different EEG bands moves towards small-world network organization during sleep.Clinical Neurophysiology,119
(9),2026–2036.Friston,K.J.,Frith,C.D.,Liddle,P.F.,&Frackowiak,R.S.(1993).Functional connectivity:The principal-component analysis of large(PET)data sets.Journal of Cerebral Blood Flow&Metabolism,13
(1),5–14.Han,S.H.,He,X.,&Woods,D.L.(2000).Hierarchical processing and level-repetition effect as indexed by early brain potentials.Psychophysiology,37
(6),817–830.Han,S.H.,Weaver,J.A.,Murray,S.O.,Kang,X.J,Yund,E.W.,&Woods,D.L.(2002).Hemispheric asymmetry in global/local processing:Effects of stimulus position and spatial frequency.Neuroimage,17
(3),1290–1299.Harness,A.,Jacot,L.,Scherf,S.,White,A.,&Warnick,J.E.(2008).Sex differences in working memory.Psychological Reports,103
(1),214–218.He,Y.,Chen,Z.,&Evans,A.(2008).Structural insights into aberrant topological patterns of large-scale cortical networks in Alzheimer's Disease.Journal of Neuroscience,28
(18),4756–4766.Herlitz,A.,Nilsson,L.G.,&Backman,L.(1997).Gender differences in episodic memory.Memory&Cognition,25
(6),801–811.Jensen,A.R.(2006).Clocking the mind:Mental chronometry and individual differences.Amsterdam:Elsevier
.Kaiser,A.,Kuenzli,E.,Zappatore,D.,&Nitsch,C.(2007).On females'lateral and males'bilateral activation during language production:A fMRI study.International Journal of Psychophysiology,63
(2),192–198.Kaiser,M.,&Hilgetag,C.C.(2006).Nonoptimal component placement,but short processing paths,due to long-distance projections in neural systems.Plos Computational Biology,2
(7),e95.Kok,A.(2001).On the utility of P3 amplitude as a measure of processing capacity.Psychophysiology,38
(3),557–577.Kosslyn,S.M.(1994).Image and brain:The resolution of the imagery debate
.Cambridge:MIT Press.Lago-Fernandez,L.F.,Huerta,R.,Corbacho,F.,&Siguenza,J.A.(2000).Fast response and temporal coherent oscillations in small-world networks.Physical Review Letters,84
(12),2758–2761.Latora,V.,&Marchiori,M.(2001).Efficient behavior of small-world networks.Physical Review Letters,87
(19),198701.Liu,Y.,Liang,M.,Zhou,Y.,He,Y.,Hao,Y.H.,Song,M.,...Jiang,T.Z.(2008).Disrupted small-world networks in schizophrenia.Brain,131
,945–961.Micheloyannis,S.,Pachou,E.,Stam,C.J.,Breakspear,M.,Bitsios,P.,Vourkas,M.,...Zervakis,M.(2006a).Small-world networks and disturbed functional connectivity in schizophrenia.Schizophrenia Research,87
(1-3),60–66.Micheloyannis,S.,Pachou,E.,Stam,C.J.,Vourkas,M.,Erimaki,S.,&Tsirka,V.(2006b).Using graph theoretical analysis of multi channel EEG to evaluate the neural efficiency hypothesis.Neuroscience Letters, 402
(3),273–277.Navon,D.(1977).Forest before trees:The precedence of global features in visual processing.Cognitive Psychology,9
,353–383.Nittono,H.,Suehiro,M.,& Hori,T.(2002).Word imageability and N400 in an incidental memory paradigm.International Journal of Psychophysiology, 44
(3),219–229.Peters,M.,&Servos,P.(1989).Performance of subgroups of left-handers and right-handers.Canadian Journal of Experimental Psychology,43
(3),341–358.Roalf,D.,Lowery,N.,&Turetsky,B.I.(2006).Behavioral and physiological findings of gender differences in global-local visual processing.Brain and Cognition,60
(1),32–42.Rubinov,M.,Knock,S.A.,Stam,C.J.,Micheloyannis,S.,Harris,A.W.,Williams,L.M.,&Breakspear,M.(2009).Small-world properties ofnonlinearbrain activity in schizophrenia.Human Brain Mapping,30
(2),403–416.Sporns,O.,&Zwi,J.D.(2004).The small world of the cerebral cortex.Neuroinformatics,2
(2),145–162.Stam,C.J.,Jones,B.F.,Nolte,G.,Breakspear,M.,&Scheltens,P.(2007).Small-world networks and functional connectivity in Alzheimer's disease.Cerebral Cortex,17
(1),92–99.Sui,D.N.,Tang,Y.Y.,Qiu,X.,Yu,Q.B.,Wang,J.H.,Zhou,L.,&Wang W.(2007).The generation of global and local mental imagery and its gender differences.Progress in Modern Biomedicine,7
,1554–1556.[隋丹妮,唐一源,邱香,于庆宝,王俊红,周莉,王威.(2007).整体表象与局部表象的产生及其性别差异.现代生物医学进展,7
,1554–1556.]Supekar,K.,Menon,V.,Rubin,D.,Musen,M.,&Greicius,M.D.(2008).Network analysis of intrinsic functional brain connectivity in Alzheimer's disease.PLoS Computationa Biology,4
(6),e1000100.Tomasi,D.,&Volkow,N.D.(2012).Laterality patterns of brain functional connectivity:Gender effects.Cereb Cortex,22
(6),1455–1462.Tononi,G.,Sporns,O.,&Edelman,G.M.(1994).A measure for brain complexity:Relating functional segregation and integration in the nervous system.Proceedings of the National Academy of Sciences United States of America,91
(11),5033–5037.Voyer,D.,&Hou,J.(2006).Type of items and the magnitude ofgenderdifferences on the MentalRotationsTest.Canadinan Journal of Experimental Psychology,60
(2),91–100.Wang,L.,Li,Y.F.,Metzak,P.,He,Y.,&Woodward,T.S.(2010).Age-related changes in topological patterns of large-scale brain functionalnetworks during memory encoding and recognition.Neuroimage,50
(3),862–872.Watts,D.J.,&Strogatz,S.H.(1998).Collective dynamics of'small-world'networks.Nature,393
(6684),440–442.Wu,J.H.,Mai,X.Q.,Chan,C.C.,Zheng,Y.Q.,&Luo,Y.J.(2006).Event-related potentials during mental imagery of animal sounds.Psychophysiology,43
(6),592–597.