4种红树净光合速率对光和CO2浓度的响应特征
2013-01-13黄丽王德炉谭芳林黄石德黄雍容
黄丽,王德炉,谭芳林,黄石德,黄雍容
(1.福建省林业科学研究院,福建 福州350012;2.贵州大学林学院,贵州 贵阳550025)
红树林(Mangrove forest)为自然分布于热带、亚热带海岸潮间带的木本植物群落,对热带、亚热带海湾生态系统的维持与发展起到关键性作用。但由于过度开发利用,红树林已大面积消失和衰退,红树林生态系统遭到了严重的破坏,这些问题已引起世界及我国政府和有关学者的极大关注。红树林生态系统的恢复现已成为系统生态恢复的一项重要内容。树种选择、造林措施是保证红树恢复成功的前提条件。树种选择、造林措施的制定跟树种的生物特性有关,而光合作用是植物最为重要的生命特征。但目前国内外对红树林光合方面尤其是自然状态下其光合作用的研究较少[1-5]。本文对秋茄(Kandelia candel)、桐花树(Aegiceras corniculatum)、白骨壤(Avicennia marina)和木榄(Bruguiera gymnorrhiza)等4种红树的光合响应曲线进行研究分析,以确定不同品种的光合能力指标,如光补偿点、光饱和点及呼吸速率等,为红树林的造林实践提供基本的理论依据。
1 材料与方法
1.1 研究区概况
福建漳江口红树林国家级自然保护区(23°53′—23°58′N,117°23′—117°32′E)属亚热带海洋性季风气候,气候温暖湿润,光、热、水资源丰富,是福建省迄今为止种类最多的红树林天然群落,也是我国北回归线以北种类最多、生长最好的红树林天然群落。该保护区年均气温21.2℃,最高月均温28.9℃(8月),最低月均温13.5℃(1月);年平均降雨量为1 714.5mm,降雨量主要集中在4—9月。保护区受降雨、江河径流和潮汐的影响,海水盐度在12‰~26‰之间变化;该海域潮汐属不正规半日潮,平均涨潮历时397min,平均落潮历时315min[6]。
1.2 试验材料
在福建漳江口红树林国家级自然保护区竹塔村附近的天然红树林同一群落内,设置样地,选取生长良好且无病虫害的秋茄、白骨壤、桐花树、木榄的平均木作为样树,每种树种各选取3株,定点观测。
1.3 研究方法
在2010年1月15日至2月10日,选择晴朗无云的早晨9:00—11:00进行光合响应曲线的测定。每种红树测定3株,每株测定1片树木冠层上部成熟、长势良好的健康叶片,结果取其平均值。测定光合响应曲线前,将待测叶在红蓝光源提供的800 μmol·m-2·s-1光强下诱导5~15min;测定时,设定温度为20℃,空气相对湿度为50%~70%;光合光响应(Pn-PAR)曲线的测定用LI-6400-02B红蓝光源设定叶室中光合有效辐射强度分别为:(2 000,)(1 800,)(1 500,)1 200,1 000,800,600,400,200,150,100,50,20,0μmol·m-2·s-1,测定红树叶片对不同光合有效辐射的净光合速率,设定CO2浓度为400μmol CO2·mol-1;光合 CO2(Pn—Ci)响应曲线的测定,以LI-6400-01液态CO2小钢瓶为气源,通过CO2注入系统控制叶室中CO2浓度分别为 1 500、1 200、1 000、800、600、400、300、200、100、50μmolCO2·mol-1,并通过LI-6400-02B红蓝光源提供饱和光合有效辐射,测定饱和光合有效辐射下,红树叶片在不同CO2浓度下的净光合速率;测定光合响应曲线时,在每个光强或CO2浓度下平衡60~200s后测定Pn值,当测量变异率小于0.05时由红外气体分析仪自动记录。
1.4 数据处理分析
应用Excel2003和SPSS13.0统计分析软件进行作图与曲线拟合。Pn-PAR响应曲线用公式Nonrectangular hyperbola(非直角双曲线)[7]和直线回归进行拟合,求得光补偿点、光饱和点、最大净光合速率、表观量子效率、暗呼吸速率;Pn—Ci响应曲线用直线回归拟合,得到CO2补偿点、羧化效率、光呼吸速率。Nonrectangular hyperbola公式如下:
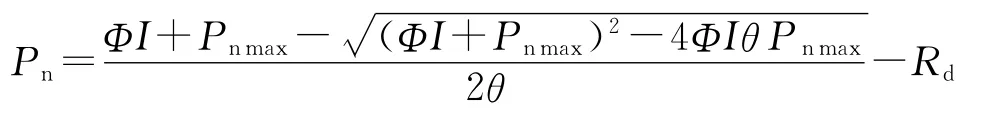
式中:Pn—净光合速率;Pnmax—最大净光合速率;Φ—表观量子效率;θ—曲角;Rd—暗呼吸速率;I—光照强度
2 结果与分析
2.1 4种红树光合作用对光强变化的响应
由于每个树种对光强的耐受程度不同,因此所设定的最高光强不同。本文中测定光响应曲线每个树种选定的最大光强是根据测定时的多次预实验而确定的。4种红树对光合有效辐射的响应曲线基本一致(图1),在PAR为0~400μmol·m-2·s-1的低光强区,光合速率随光强的增加而成比例地增加,在高光强区,光合速率增加明显变缓,当超过一定的光强时,光合速率就几乎不再增加。
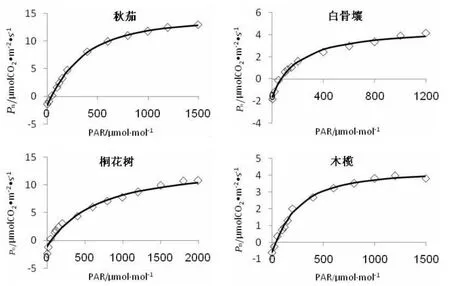
图1 不同红树叶片Pn对光响应曲线
不同红树光饱和点(LSP)和光补偿点(LCP)不同(表1)。光补偿点在39~81μmol·m-2·s-1之间,白骨壤>桐花树>秋茄>木榄。利用公式Nonrectangular hyperbola(非直角双曲线)[7]求得的光饱和点在450~650μmol·m-2·s-1之间,虽然跟试验观测值差异较大,明显比图1中的试验观测值小,但还是能客观反映各树种光饱和点的差异,因此以下分析仍使用此值。光饱和点从大到小顺序排列为:桐花树>秋茄>白骨壤>木榄。
表观量子效率(AQY)是叶片光能利用效率的一个重要指标,反应叶片对光尤其是弱光的利用能力[8],不同红树间表观量子效率存在差异,秋茄的表观量子效率最高,木榄最低,说明秋茄叶片的光能利用效率最大,对弱光的利用能力最大,木榄最小;暗呼吸速率(Rd)与叶片的生理活性有关,桐花树暗呼吸速率最大,秋茄次之,白骨壤稍低于秋茄,木榄最低(表1)。
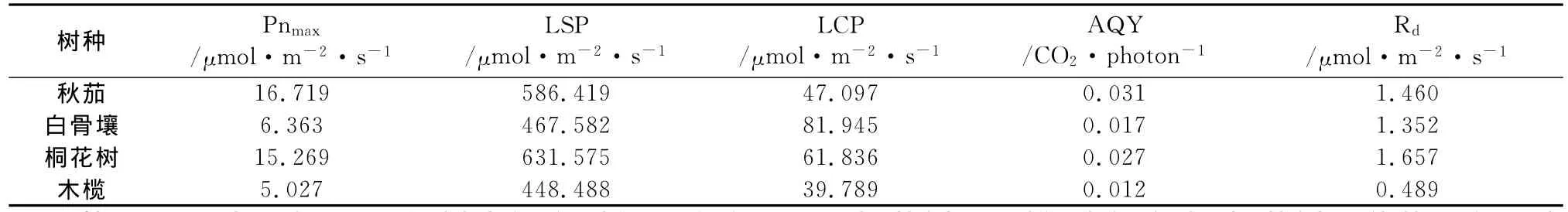
表1 不同红树的光响应参数
从图1可以看出,不同红树光饱和时的最大净光合速率大小排列顺序,与曲线拟合所得的最大光合速率(Pnmax)结果一致(表1),拟合结果为:秋茄>桐花树>白骨壤>木榄。光饱和时的Pnmax,可以反映叶片的光合潜能。以上结果表明,秋茄和桐花树的光合潜能明显高于白骨壤与木榄。
2.2 4种红树光合作用对CO2浓度变化的响应
Pn-Ci响应曲线拟合参数计算结果见表2。4种红树光合作用的CO2补偿点在103~152 μmolCO2·mol-1之间,远 大于 30μmolCO2·mol-1,说明这4种红树均为 C3植物[9],木榄 CO2补偿点最高,白骨壤次之,桐花树和秋茄较低。4种红树的光呼吸速率在4~9μmolCO2·mol-1之间,桐花树最高,秋茄次之,白骨壤、木榄比较接近。4种红树羧化效率(CE)从小到大排列顺序与光呼吸一致,最大的为桐花树,第2为秋茄,第3为白骨壤,木榄最小。
3 结论与讨论
由于光合作用与植物生长地的环境条件有密切关系,光合作用的某些生理参数,如Pnmax、LSP、LCP等,已成为植物制定栽培措施的科学依据[10],植物叶片的光饱和点和光补偿点反映了植物对光照条件的要求,一般情况下光饱和点和光补偿点较低的植物属于耐阴植物,反之属于阳性植物。光饱和点较高和光补偿点较低的植物对光环境的适应性较强,反之则较弱。4种红树光补偿点,白骨壤>秋茄>桐花树>木榄,光饱和点,桐花树>秋茄>白骨壤>木榄。光饱和点及光补偿点的差异,说明不同红树树种之间可利用的光强范围及对高低光强的适应能力存在差异,因此在进行红树造林时,要注意树种的配置及造林措施,可以先种光饱和点较高、对强光的适应性高的种类,以后再套种光饱和点低、耐阴一点的种类。桐花树的光饱和点最高,说明其对强光的适应能力较强,这可能跟桐花树叶片的泌盐特性高有关,叶片泌盐不仅可以把桐花树体内多余的盐分排出体外,同时叶片上的盐分也可以把多余的光线反射出一部分,从而避免强光对叶片光合机构的破坏,而保持较高的光合能力,在实验中也发现,中午桐花树光合受到光强胁迫的程度小于其他树种。
4种红树光合作用的CO2补偿点在103~152 μmolCO2·mol-1之间,与湿地松的CO2补偿点(163 μmolCO2·mol-1)[11]、杉木的CO2补偿点(130.4~163.5μmolCO2·mol-1)[12]以及一些阔叶树种的CO2补偿点,如银杏(100μmolCO2·mol-1)[13]、黄背栎(105.21μmolCO2·mol-1)[14]等一些陆生树种的补偿点相比,相差不大。4种红树羧化效率(CE),桐花树为0.0705,秋茄为0.064,白骨壤为0.036,木榄为0.027,均高于湿地松的CE(0.017)[11],除木榄外其他树种均高于黄背栎的CE(0.035)[14]。红树这种光合特性,可能跟长期适应其生存的逆境环境有关,CO2补偿点相当,但CE却相对较高可以使红树植物在相同的CO2浓度(不对光合能力产生抑制)条件下,比其他普通树种生产出更多维持其在水淹逆境环境中生存的产物,因为植物在淹水胁迫下产生能量效率低的糖酵解代谢、乙醇发酵和乳酸途径,这些代谢途径须消耗体内大量贮存的碳水化合物,才可产生足够的ATP,维持细胞的生存和功能[15]。
[1]杨盛昌,中须贺常.夜间低温对红树幼苗光合特性的影响[J].厦门大学学报:自然科学版,1999,38(4):617-622
[2]杨盛昌,中须贺常雄,林鹏.5℃夜间低温对红树幼苗光合速率和蒸腾速率的影响[J].植物研究,2001,21(4):587-591
[3]杨盛昌,中须贺常雄,林鹏.光强对秋茄幼苗的生长和光合特性的影响[J].厦门大学学报:自然科学版,2003,42(2):242-247
[4]杨坚,张玲玲,何斌源,等.互花米草混种密度对秋茄幼苗生理生态的影响[J].福建林学院学报,2007,27(2):176-179
[5]段文芳,石贵玉,秦丽凤,等.镉胁迫对桐花树光合·蒸腾作用及保护酶活性的影响[J].安徽农业科学,2008,36(4):1355-1356,1370
[6]林鹏.福建漳江口红树林湿地自然保护区综合科学考察报告[M].厦门:厦门大学出版社,2001
[7]Farquhar G D,Sharkey T D.Stomata conductance and photosynthesis[J].Annum Review Plant Physiology,1982,33:317-345
[8]蹇洪英,邹寿青.地毯草的光合特性研究[J].广西植物,2003,23(2):181-184
[9]许大全.光合作用效率[M].上海:上海科学技术出版社,2002:119-119
[10]朱万泽,王金锡,薛建辉.台湾桤木引种的光合生理特性研究[J].西北植物学报,2004,24(11):2012-2019
[11]谭芳林,薛建辉,张水松,等.不同混交林中湿地松的光合特性[J].南京林业大学学报:自然科学版,2003,27(4):35-37
[12]俞新妥.不同种源杉木光合特性的比较研究[J].福建林学院学报,1989,9(3):223-227
[13]孟庆伟,王春霞,赵世杰.银杏光合特性的研究[J].林业科学,1995,3(1):69-76
[14]朱万泽,吴永波,薛建辉.贡嘎山地区黄背栎的光合特性[J].南京林业大学学报:自然科学版,2006,30(1):25-28
[15]李阳生,王建波.淹涝胁迫对水稻叶鞘和叶片中淀粉粒分布的影响[J].武汉植物学研究,2000,18(6):528-530
