长白山杨桦次生林树种空间隔离度对林分生长的关系
2012-12-29亢新刚谢佳利黄新峰冯启祥
孔 雷 ,亢新刚 ,杨 华 ,谢佳利 ,黄新峰 ,冯启祥
长白山杨桦次生林树种空间隔离度对林分生长的关系
孔 雷1,亢新刚1,杨 华1,谢佳利1,黄新峰2,冯启祥3
(1.北京林业大学 林学院 省部共建森林培育与保护教育部重点实验室,北京 100083;2. 河南省林业调查规划院,河南 郑州 450045;3.汪清林业局,吉林 汪清 133200)
以6块立地条件、林分密度和混交比相同的长白山天然杨桦次生林固定样地为研究对象,样地内每木检尺和定位。通过系统聚类方法划分出树种多样性和物种均匀度相同的样地,利用树种多样性混交度研究林分蓄积生长量与物种空间隔离度的关系。结果表明:在立地条件、株数密度、混交比和树种多样性相同时,样地内林分蓄积生长量与树种空间隔离度呈反比关系。最后给出树种配置方法的建议,为今后经营合理的天然次生林空间结构给予启示。
树种多样性混交度;林分蓄积生长量;树种多样性;物种均匀度;株数密度;混交比
林分结构是天然林经营管理重要内容,营造良好的天然林林分结构能够极大促进林分的生长发育和持续[1-2]。林分结构分为空间结构和非空间结构。林分空间结构分为林木空间分布格局,竞争关系,物种空间格离度。传统方法是用混交比[3]、Shannon-wiener多 样 性 指 数[4]、Simpson生态优势度指数[5]、Pielou均匀度指数[6]等来表示树种的配置情况,但这些指数都是非空间结构指数,还不能说明树种空间分布情况。Clark和Erans[7]通过利用对象木与最邻近树木特征的比较,来描述树种之间空间配置关系,并且称之为树种空间格离度。Gadow等[8]提出用混交度方法来表示树种空间格离度,但是这种方法仅比较对象木与最近邻木之间树种异同,而没有考虑最近邻木相互之间的树种异同。鉴于此汤孟平[9]提出了能够量化描述最近邻木之间空间隔离度的树种多样性混交度。
因为次生林在演替过程中具有较高的树种多样性[10],包含顶级耐萌性树种和先锋树种,所以利用树种多样性混交度研究杨桦次生林物种在林分中的配置情况具有实际意义。杨桦次生林是阔叶红松林被破坏后的次生裸地上首先发生的先锋群落,是阔叶红松林自然恢复演替规律的重要过渡阶段之一[11]。杨桦次生林的优势树种主要有:山杨、白桦和枫桦3种。目前,国内对杨桦次生林的非空间结构研究主要是直径结构和年龄结构。龚直文[12]通过weibull分布和负指数分布模拟和检验了长白山区杨桦次生林的直径结构。罗菊春[13]从年龄结构分析了白桦次生林的生产力和非空间结构特点。对杨桦次生林的空间结构研究主要是空间分布格局和空间物种格离度。龚直文[11]运用点格局方法分析了杨桦次生林长白山区森林演替过程中的特点和发展方向。孔令红[14]利用角尺度研究长白山区山杨次生林的空间分布格局。这些研究有助于了解次生林林分结构和林分功能,但是何种树种配置结构是最优的结构,怎样才能够最大地发挥天然次生林林分功能还没有相关研究。本文基于树种多样性混交度的方法,从树种空间格离度的角度对长白山区杨桦次生林进行分析,深度研究长白山区杨桦次生林的生长与树种空间结构的关系,为提高长白山区杨桦次生林的林分蓄积生长量和经营管理水平和使之趋向于理想林分结构状态提供理论和实践依据。
1 试验地概况
试验研究地区位于吉林省东部长白山北麓的金沟岭实验林场,东经129°56′~130°04′,北纬43°05′~ 48°40′之间,海拔 300 ~ 1 200 m。该地区属于温带大陆性季风气候,年平均气温3.9 ℃,极端最高气温37.5 ℃,极端最低气温-37.5 ℃。年温度大于10 ℃的活动积温为2 473 ℃,年平均日照时数约为2 358 h,年降水量均在500~600 mm。初霜从9月中旬开始,终霜期一般在5月中旬,积雪日数为80~100 d。风向的季节性变化明显,风速为2~5 m/s。研究数据来源于金沟岭实验林场杨桦次生林的6块立地条件和林分密度相同的固定样地。优势树种有白桦,枫桦和山杨。其他树种有:椴木、水曲柳、蒙古栎鱼鳞云杉、山槐、色木槭、红松和臭冷杉。样地面积为0.2 hm2,样地林木株树为278~356株。该样地从2007年至2011年每2年连续调查一次,如表1所示。调查内容:对胸径(DBH)大于5 cm的树木进行每木检尺,采用全站仪定位,测定每株树木坐标(X,Y),并记录树种名称,树高,冠幅,第一活枝下高。
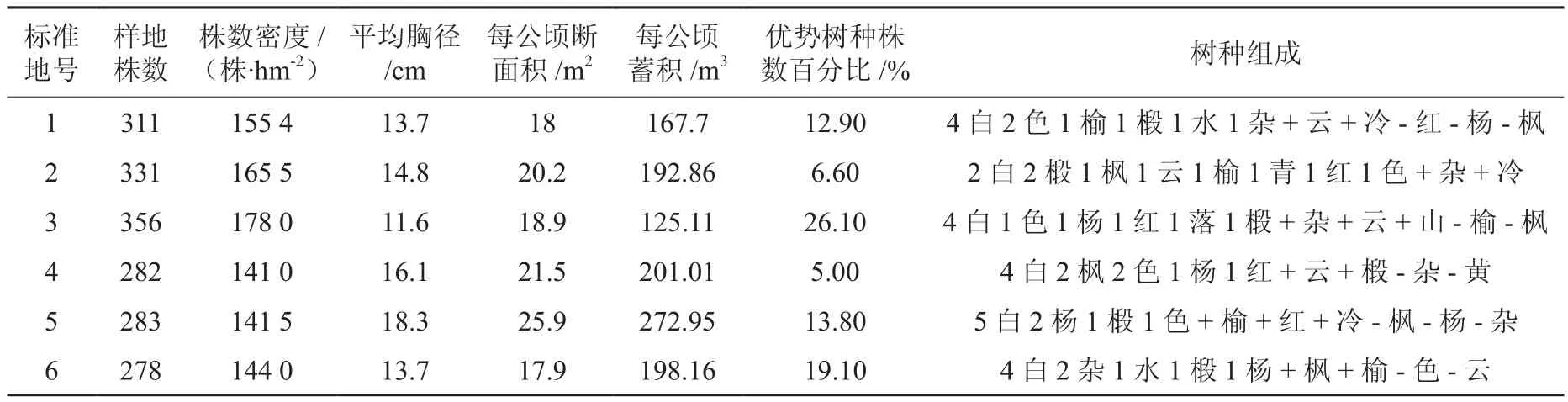
表1 标准地的基本概况Table 1 The basal statistics of samples
2 研究方法
2.1 树种多样性混交度
树种多样性混交度公式为:

式中:Mi是林木树种多样性混交度;ni为目标树邻近木之间不同的树种个数;n为邻近木株树。当n取值为4时,相邻木与目标树构成5种结构关系,即零度、弱度、中度、强度、极强度过渡阶段完整,生物学意义明显,空间结构信息完整[11],本研究采用n=4的树种多样性混交度进行分析。
2.2 系统聚类法
常用的聚类方法有系统聚类法、动态聚类法、有序样品聚类法和图论聚类法。这些方法归类步骤一致,类间距离计算方法不同[16-17],本研究采用离差平方和法对林分株树密度进行聚类分析。离差平方和法是由ward提出来的,其思想源于方差分析:如果分类正确,同类样品的离差平方和法事小,类与类之间的离差平方和应当较大[18]。
设将n个样品分成 k类,G1,G2,…,Gk,Xit表示类Gt中第i个样品,nt表示类Gt中样品个数,Xt是类Gt的重心,则在类比Gt中样品离差平方和是:

如果Gp和Gq合并成新类Gr,则类内离差平方和分别为:
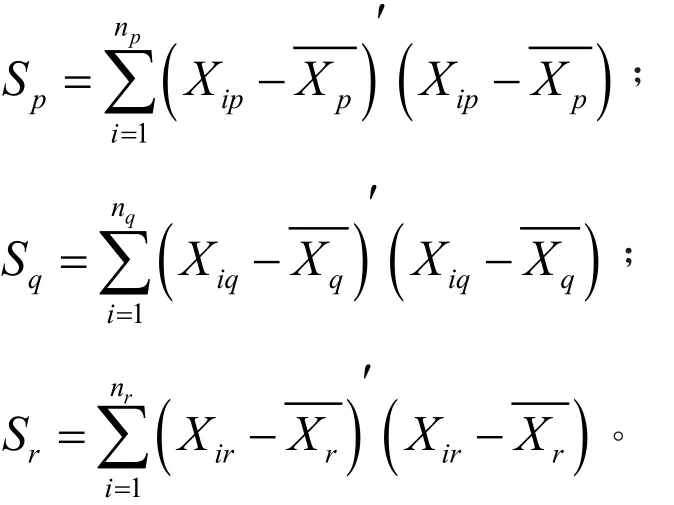
定义Gp和Gq之间平方和距离为:

式中: Gr= Gp∪ Gq。则类Gk与新类Gr的距离递推公式:
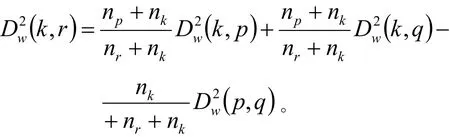
2.3 边缘校正
当对象木到样地边界距离小于到基邻近木距离时,必须进行边缘校正[19]。目前,在校正边缘效应时,常用的方法有加权校正法,平移式八邻域大样地边缘校正法和缓冲区校正法等。加权校正法假设研究区域内外事件分布特征相似,通过研究区域内的事件数来估计研究区域外事件数从而校正边缘效应。平移式八邻域大样地边缘校正法是在原样地的上、下、左、右、左上、右上、左下、右下,8个方向复制原样地,形成由9个相同样地组合的大样地,然后计算原样地的混交度。这两种方法有一个共同的缺点就是为了提高估计结果无偏性,增加对研究区域外事件数的估计。而缓冲区校正法则没有这样的局限,其校正原理是通过缩小标准样地面积的方法排除边缘影响,从边界开始向内减少长度。比较不同空间尺度对象木之间空间影响,直到影响不在变化或很小时得到缓冲区的长度。这种方法也有其不足,就是缓冲区研究事件相交信息没有充分被利用,而且没有确定的标准来指定缓冲区的长度,缓冲区长度太小仍有边缘效应,太大则大量有用信息被遗漏[20]。本文采用缓冲区校正法通过扩大样地面积,增加样地数量来弥补该方法的缺陷。
2.4 树种多样性
对于树种多样性的测定,本文采用下式计算:
式中:S为样地物种数,Ni为第i个种的个体数,N为样地总个体数。
3 结果与分析
3.1 树种多样性和均匀度
在立地条件和林分密度基本相同情况下,计算各样地的树种多样性。杨桦次生林的优势树种是桦树和杨树,属于强阳性先锋树种,需要占有大量光照,一旦处于被压状态不能正常生长。在样地2中,优势树种株数占总株数偏小的样地,种间竞争相对较弱,一些耐荫树种如红松、臭冷杉、鱼鳞云杉、红皮云杉等能够在这些强阳性先锋树种下正常生长,充分利用各林层的营养空间,从而使林分密度增大,树种多样性变多,林分总生长量增大。反之则树种多样性变小,林分总生长量减少(表2)。杨树和桦树具有较强的根萌分蘖能力和天然更新能力,降低了其他种群的物种毗邻程度。由于这种繁殖方式,如果杨树和桦树株数比重较大,则使样地均匀度降低,呈现聚集分布状态(表2)。
3.2 树种多样性聚集分析
表2可得树种多样性和均匀度对林分生长的影响是较明显的,应该分成不同等级下研究树种空间隔离度对林分生长的影响。利用系统聚类分析方法,根据样地Shannon-wiener多样性指数指数H将6块样地分为3类。聚类结果见图1和表3。样地6为树种多样性低林分,样地1、3、4和5为树种多样性中林分,样地2为树种多样性高林分。
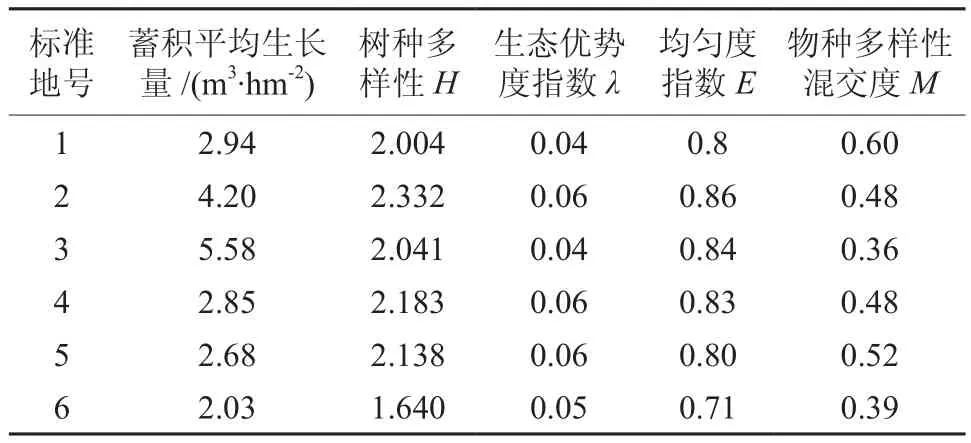
表2 各样地的林分生长量、树种多样性、生态优势度指数、均匀度指数和物种多样性混交度Table 2 Stand volume increment, tree species diversity,dominance, evenness and diversity mingling indices of plots

图1 物种多样性系统聚类的树形图Fig.1 Dendrogram of tree species diversity

表3 物种多样性系统聚类Table 3 Agglomeration of tree species diversity
3.3 树种空间隔离度对林分生长量的影响
在林分形成初期,桦树和杨树以萌芽和种子更新的方式繁殖,这种强繁殖方式加剧了种内竞争,淘汰生长不良或有病虫害的林木,保存生长力旺盛的林木,从而形成不同树种间穿插程度降低,树种空间隔离度较小的情况。桦树和杨树单位面积株数越多,通过种内竞争得到长势优良林木的质和量就越高。由于优势树种生长很大程度上决定了林分蓄积生长量的大小,因此这种林分生长量较高(表2)。相反杨树和桦树单位面积株数越少的林分,保留的林木质和量次之。虽然这在一定程度上削弱了种间竞争,给了一些耐荫树种生长的空间,而且增加了林分物种的多样性和均匀度,但是耐荫树种在幼期生长缓慢,总林分生长量上还是较低。所以杨桦次生林生长量较大的林分特征体现为树种多样性较低、空间隔离度低、聚集的分布状态。在选择的1、3、4和5样地立地条件、林分密度和树种多样性都基本相同条件下,这些样地林分生长量与树种空间隔离度呈反比关系(图2)。

图2 林分生长量与物种空间隔离度的关系Fig. 2 Relationship of stand volume increscent and tree species diversity mingling
3.4 不同生活史树种空间隔离度对林分生长量的影响
如图2所示3号样地林分蓄积生长量最高,3号样地上层白桦和杨树等强阳性树种居多,下层冷杉等耐荫性树种居多。当上层先锋树种达到过熟枯死后,下层耐荫树种就会取代上层先锋树种,林分空间格局由聚集状态演变为均匀状态,而树种空间隔离度则由小变大,表明树种间穿插程度较高。由此可见这种林分结构在向顶级群落演替过程中能够保持较大的林分蓄积生长量。如表4所示。

表4 样地1、3、4和5号不同径阶胸径的树种丰富度、多样性、均匀度和物种多样性混交度Table 4 Tree species richness, diversity, evenness and diversity mingling indices in different DBH classes of plot 1, 3, 4 and 5
4 结论与讨论
在经营杨桦次生林时,既不能忽略树种的生物学特性,营造高混交状态,也不能忽略种间竞争任其繁殖和发展。研究物种空间隔离度对林分生长量的影响时由主到次的顺序初步需要确定5个因子:首先要考虑天然杨桦次生林的立地质量。科学评定天然林地生产潜力。结合气候因子、土壤因子和生物因子,生产适宜该地区生境的树种。同时充分考虑林木生物学特性,促进林木合理生长。本区低山灰化灰棕壤土是适宜杨桦次生林生长的立地类型。其次林分密度对林分蓄积生长的影响也是很大的。密度低的林分单株材积大,蓄积生长量大,但单位面积株数少,密度大的林分单位面积株数多,但单株材积少,蓄积生长量小,密度中等的林分单位面积和蓄积生长量是最大的[21]。本研究是在这项研究的基础上进一步研究物种空间隔离对林分生长量的影响。兰士波[21]指出林分密度在1 600株/hm2左右,林木的平均干物质积累和光能利用率最大,同时营养元素的归还量和单位面积上的胸径年生长量都是最大的。研究样地株数密度在1 500株/hm2左右,接近这一株数密度。再次,林分树种的混交比也是影响因子之一。在研究林分空间结构时,没有考虑林分的混交比,而直接研究林分的混交度是没有科学意义的[22]。最后,树种多样性表示某一树种在样地内出现的频率。树种多样性的不同,林分生长情况必然也不同。树种多样性能够反映群落结构类型,组织水平、发展阶段、稳定程度和生境差异等,所以在研究林分生长与树种空间隔离度关系时,树种多样性也不能忽略[23]。本研究将树种多样性聚类分析,提取相同类的样地做进一步的分析,最后得到物种空间隔离度与林分生长量成反比关系。在林分生长初期,聚集状态的林分空间结构更有利于优势树种的生长,经过一段时期种内竞争,存活下来一定数量优良林木,然后在乔木下层更新耐荫树种,逐渐营造均匀状态的林分空间格局,从而实现对林分蓄积的优化经营。
[1] 方国景,汤孟平,章雪莲. 天目山常绿阔叶林的混交度研究[J]. 浙江林学院学报, 2008, 25(2): 216-220.
[2] 吕 勇,李卫兵,王才喜,等. 黄丰桥林场杉木林分直径的结构特征[J]. 中南林学院学报,2002, 22(2): 44-47.
[3] Graz F P. The behavior of the species mingling index Msp in relation to species dominance and dispersion[J]. Eur. J. Forest Res., 2004, 123(1): 87-92.
[4] 肖 益,田大伦,闫文德,等. 沅陵3种天然次生林群落乔木层物种多样性研究[J]. 中南林业科技大学学报, 2011,31(5): 121-126.
[5] 李际平,郑 柳,赵春燕,等. 不同等级廊道杉阔林下植被物种的多样性分析[J]. 中南林业科技大学学报, 2012, 32(2):64-69.
[6] 蔡 聪. 马尾松阔叶树混交林物种多样性的初步研究[J]. 中南林学院学报, 1995, 15(2): 195-200.
[7] Clark P J, Evans F C. Distance to nearest neighbor as a measure of spatial relationships in populations[J]. Ecology, 1954, 35(4):445-453.
[8] Gadow V K, Fueldner K. Zur Methodik der Bestand beschreibung[R]. Klieken: Vortrag Anlaesslich der Jahrestagung der A G Forsteinrichtung, 1992.
[9] 汤孟平,唐守正,雷相东,等. 两种混交度的比较分析[J].林业资源管理, 2004, 8(4): 25-27.
[10] Connell J H. Diversity in tropical rain forests and coral reefs[J].Science, 1978, 199: 1302-1310.
[11] 龚直文,亢新刚,顾 丽,等. 长白山杨桦次生林生长过程与演替动向分析[J]. 林业科学研究, 2009, 22(3): 379-384.
[12] 龚直文,亢新刚,杨 华,等. 长白山杨桦次生林直径结构研究[J]. 西北林学院学报, 2009, 24(3): 1-6.
[13] 罗菊春. 北京百花山白桦次生林的结构与生产力[J]. 北京林学院学报, 1984(4): 8-19.
[14] 孔令红,郑小贤. 金沟岭林场次生林空间分布格局分析[J].林业调查规划, 2007(4): 3-7.
[15] 林勇明,洪 滔,吴承祯,等. 桂花次生林群落主要树种种间关联及其对混交度的响应[J]. 应用与环境生物学报, 2007,13(3): 327-332.
[16] 朱建平. 应用多元统计分析[M]. 北京: 科学出版社, 2006.
[17] 余加林,肖枝洪. 多元统计及SA S应用[M]. 武汉: 武汉大学出版社, 2008.
[18] 何晓群. 多元统计分析[M]. 北京: 中国人民出版社, 2004.
[19] 吴巩胜,王政权. 水曲柳落叶松人工混交林中树木个体生长的竞争效应模型[J]. 应用生态学报, 2000, 11(5): 646-650.
[20] Diggle P. statistical analysis of spatial point pattens[M]. 2 ed.London: Edward Arnoid publisher, 2003.
[21] 兰士波. 天然杨桦林密度效应的研究[J]. 南京林业大学学报:自然科学版, 2007, 31(2): 83-87.
[22] 惠刚盈,Gadow K V,胡艳波. 结构化森林经营 [M]. 北京:中国林业出版社, 2007.
[23] 钱迎倩,马克平. 生物多样性的原理和方法[M]. 北京: 中国科学技术出版社, 1994.
Relationship between spatial isolation of trees and stand volume increment of polar-birch secondary forests in Changbai mountain
KONG Lei1, KANG Xin-gang1, YANG Hua1, XIE Jia-li1, HUANG Xin-feng2, FENG Qi-xiang3
(1.Key Lab. for Silviculture and Conservation of Ministry of Education, College of Forestry, Beijing Forestry University, Beijing 100083, China; 2. Henan Provincial Academy of Forest Inventory and Planning, Zhengzhou 450045, Henan, China; 3. Wangqing Forest Bureau, Wangqing 133200, Jilin, China)
The study was based on 6 permanent plots data of polar-birch secondary forests in Changbai Mountain, which owned the same site quality, mixed proportion and diversity of trees. The plots were located and the tree diameters at breast heights were investigated. By applying hierarchical cluster method, the plots were classified according to diversity of trees and species evenness. Then the relationship between stand volume increment and spatial isolation of trees was analyzed by tree species diversity mingling method. With the same of site quality, diversity of trees, diversity of tree and mixed proportion, the results manifested that the stand volume increment in the plots declined with the incensement of spatial isolation of trees. Finally, the reasonable advices of trees spatial isolation were given, which helps to manage an optimal spatial structure.
tree species diversity mingling; stand volume increment; tree species diversity; species evenness; strains density; mixed proportion
S757.2
A
1673-923X (2012)07-0014-05
2012-03-23
林业公益行业项目(200804027)
孔 雷 (1986—),男,山东曲阜人,博士研究生,主要从事天然林林分结构的研究;E-mail :1234konglei4321@sina.com
亢新刚(1952—),男,北京人,教授,主要从事天然林经营管理的研究;E-mail : xingangk@163.com
[本文编校:欧阳钦]
