野生小蓬竹的光合光响应曲线及其模型拟合
2012-12-28廖小锋刘济明张东凯闫国华
廖小锋,刘济明,张东凯,靳 勇,张 勇,闫国华,王 敏
野生小蓬竹的光合光响应曲线及其模型拟合
廖小锋,刘济明,张东凯,靳 勇,张 勇,闫国华,王 敏
(贵州大学 林学院, 贵州 贵阳 550025)
应用5种典型的光响应模型对野生小蓬竹叶片光响应曲线进行了拟合,并探讨了几种模型在光响应研究中的适用性。结果表明:①直角双曲线、非直角双曲线及指数函数模型无法求取植物最大净光合速率(Pnmax)和光饱和点(Isat)的解析解,而结合其它方法拟合的相应值却与实测值相差很大,同时也不能处理光抑制部分的光响应数据;②二次多项式模型能够一定程度地处理光抑制部分的光响应数据,但其拟合得到的各项光合参数与实测值相差较大,甚至会出现逻辑错误;③修正的直角双曲线模型拟合的各项光合参数均比较准确,同时也能很好地处理发生光抑制部分的光响应数据;④采用修正的直角双曲线模型拟合的野生小蓬竹Pnmax、Isat、光补偿点(Ic)、暗呼吸速率(Rd)和初始量子效率(α)分别为 8.53 µmol·m-2s-1、1 750.75 µmol·m-2s-1、21.40 µmol·m-2s-1、1.06µmol·m-2s-1、0.054。
小蓬竹;光响应曲线;光响应模型;修正的直角双曲线
小蓬竹Drepanostachyum luodianense系竹亚科镰序竹属,其地下茎合轴型,秆柄短缩,竹秆在地面密集丛生。小蓬竹目前虽然仅分布于贵州罗甸县、平塘县、长顺县及紫云苗族布依族自治县的喀斯特山地,但其生态适应能力强大,是喀斯特适生竹种,在喀斯特生态系统的水源涵养、水土保持、养分平衡等生态功能发挥中具有重要作用[1]。光合作用是植物生长发育的基础,同时又是一个复杂的生物物理化学过程,受到环境生态因子及生理因子的共同影响。光合速率、表观量子效率、光饱和点、光补偿点等可由植物光响应曲线估算的光合参数是植物重要的生态生理参数[2],因此,确定植物光合作用的光响应曲线对于研究植物的光合特性具有重要意义[3]。1905年,Blackman提出了第一个光合作用光响应模型[4]之后,直角双曲线模型[5]、非直角双曲线模型[6]、指数函数模型[7-9]和修正的直角双曲线模型[10-11]相继提出;此外,国内应用较多的还有二次多项式模型[12-13]。本文中采用上述常用的5种模型对小蓬竹光响应数据进行了拟合,探讨了几种光响应模型的适用性,同时也为小蓬竹以后的科学栽培和管理提供依据。
1 研究地自然概况
研究地为小蓬竹典型自然分布地之一——贵州罗甸县董架乡董架村。董架村位于罗旬县西北部,该区域出露的地层以三叠系的纯质石灰岩为主,为典型喀斯特丘陵地貌景观,石灰岩基岩大面积裸露,地面崎岖不平,溶蚀洼地和峰丛分布较广,洼峰之间的相对高度均在100 m以上;气候属中亚热带季风气候,年降水量在1 300 mm左右;原生植被为亚热带常绿阔叶林,由于长期遭到人为破坏,原生森林植被多已逆向演替为落叶阔叶林、灌丛、草坡等不同类型的次生植被。试验地位于董架村,该地分布的小蓬竹群落较为典型。试验地海拔925 m,坡向为东北坡,坡位为中坡位,地理位置为东经106°54′20″、北纬25°37′17″,土壤为典型的喀斯特石灰土,水肥条件较好。小蓬竹群落内伴生植物主要为朴树Celtis sinensis、乌桕Sapium sebiferum、女贞Ligustrum lucidum、青檀Pteroceltis tatarinowii、皱叶雀梅藤Sageretia rugosa、绣线菊Spiraea salicifolia、铁仔Myrsine africana、荩草Arthraxon hispidus、淡竹叶Lophatherum gracile、白酒草Conyza japonica、五节芒Miscanthus floridulus等。
2 材料与方法
试验材料为小蓬竹分布地典型群落林中的2年生小蓬竹植株。在林中选择3丛生长良好的小蓬竹,每丛选择1株2年生平均株,每株选取3片处于林冠上层的植株上部当年生向阳成熟功能叶,标记以备测定。
2.1 光响应数据的测定
光响应数据于2011年8月初小蓬竹出笋期利用美国生产的Li-6400便携式光合作用测定系统进行测定。选择晴朗无风(微风)的天气,于早上9:00~11:30之间进行,这段时间植株最为活跃,可省去叶片活化环节。测定时,控制叶室中CO2浓度为 380±2 µmol·mol-1,空气相对湿度为(70±5)%,叶片温度控制为28±0.5℃。在控制条件下,对Li-6400的02B-LED红蓝光源进行设置,使光合有效辐射(I)从 2 000 µmol·m-2s-1逐渐降低到 0 µmol·m-2s-1(设置值分别为 2 000、1 800、1 600、1 400、1 200、800、500、300、150、100、50、20、0),采用自动程序测量,每个设置值下停留3 min。每片叶子重复测定3次,取平均值。
2.2 光响应曲线模型
2.2.1 二次多项式模型
二次多项式模型[12-13]表达式为:

式(1)中:Pn为净光合速率;I为光合有效辐射;a、b、c为系数。二次多项式在光响应曲线研究中仅作为一个纯数学模型被应用,因此,式(1)中a、b、c这3个参数都不具备生物学意义。根据二次多项式的特性可以求取光饱和点Isat、光补偿点Ic、最大净光合速率Pnmax、暗呼吸速率Rd、初始量子效率α的解析解。表观量子效率(apparent quantum efficiency,AQE,A)只能利用直线方程拟合弱光下(≤200 μmol·m-2s-1)的光响应数据得到[14]。
2.2.2 直角双曲线模型
光响应曲线的直角双曲线模型[5]表达式为:
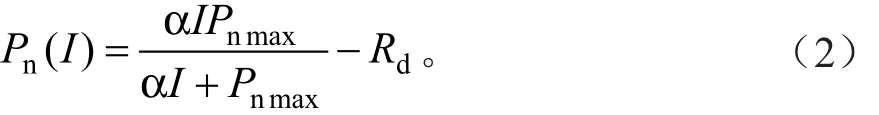
式中:Pn、I的定义同前;α为植物光合作用对光响应曲线在I=0时的斜率,即光响应曲线的初始斜率,也称为初始量子效率;Pnmax为最大净光合速率;Rd为暗呼吸速率。式(2)是一个没有极值的函数,也即直角双曲线是一条没有极点的渐近线,无法由式(2)求出植物的Pnmax和光饱和点的解析解。因此,必须利用非线性最小二乘法估算Pnmax,而对于Isat,需要求解直线方程式(3)得到[15],其中AQE求解方法同前:

2.2.3 非直角双曲线模型
光响应曲线的非直角双曲线模型[6]表达式为:
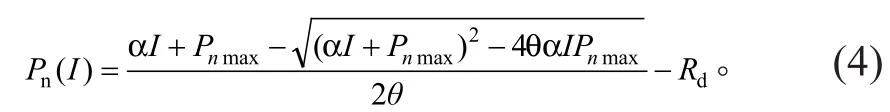
式 中:Pn、α、I、Pnmax、Rd的 定 义 同 前,θ 为反映光响应曲线弯曲程度的曲角参数,取值0≤θ≤1。式(4)同样是一个没有极值的函数,无法求取Isat的解析解,估算Isat的方法与直角双曲线模型的方法相同,AQE求解方法同前。
2.2.4 指数函数模型
光响应曲线的指数函数模型形式较多,由Bassman和Zwier在1991年首次给出,随后Prado和Moraes在1997年、Watling等在2000年也提出过相关模型。虽然指数函数模型形式较多,但它们的本质是一样的,即都是没有极值的函数。本文中选取Bassman和Zwier提出的指数函数模型[7],其表达式为:
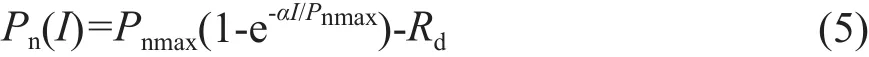
式中:Pn、α、I、Pnmax、Rd的定义同前,e为自然对数的底。通过式(5)可以求取Ic的解析解,但无法求取Isat,要估算饱和光强,需假设光合速率为0.9 Pnmax或0.99 Pnmax所对应的光强为饱和光强[16-17],AQE求解方法同前。
2.2.5 修正的直角双曲线模型
修正的直角双曲线模型是Ye和Yu提出的[10-11],其表达式为:
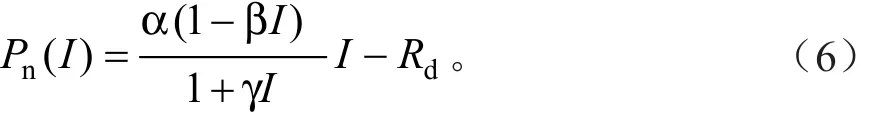
式中:α、I、Rd的定义同前,β和γ为系数。由于式(6)是一个具有极值的函数,因此,由式(6)可以直接求取Isat、Ic、Pnmax的解析解,AQE求解方法同前。
2.3 数据处理和分析
试验数据的初步处理由Excel2003完成,模型分析由SPSS17.0完成。
3 结果与分析
表1给出了野生小蓬竹光合作用光响应实测数据及上述5种模型的拟合值。由表1可知,二次多项式模型拟合值与实测值相差较大,甚至在I为0 μmol·m-2s-1时给出的Pn为正值;其余4种模型在各个数据点下的拟合值与实测值都较为接近,但仅有修正的直角双曲线模型能很好地处理小蓬竹Pn在高光强部分下降的问题。
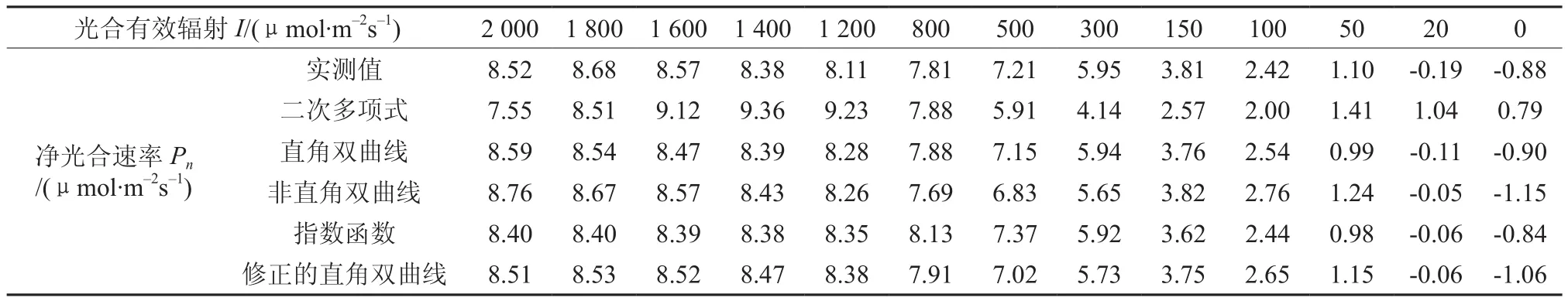
表1 野生小蓬竹光合作用光响应曲线测量数据及其拟合值Table 1 The measured and simulated data of light-response curve of photosynthesis of wild D.luodianense
表2给出了用上述5种模型分别拟合表1中野生小蓬竹光响应实测数据得到的光合参数。由表2可知,二次多项式模型拟合得到的光合参数有违背生物学常识的地方(Ic、Rd为负值);通过直角双曲线、非直角双曲线及指数函数直接拟合以及二次多项式求解得到的Pnmax与实测值相差较大;同时还应看到,二次多项式求解得到的Isat虽然较大,但与实测值仍有较大差距,而直角与非直角双曲线通过直线方程求解得到的Isat则远小于实测值,指数函数需要假设Pn为0.9Pnmax所对应的光强Isat,但同样远小于实测值,如果按0.99Pnmax进行假设则无法求取Isat。由表2可知,尽管直角双曲线、非直角双曲线与指数函数给出的决定系数R2>0.99,但它们拟合得到的光合参数与实测值相差较大,因此,R2越大只能说明模型拟合程度越高,但并不能保证拟合结果就一定与实测值相符。此外,对比不同模型所给出的光合参数,只有修正的直角双曲线求解得到的各项光合参数与实测值均较为吻合。

表2 5个模型拟合得到的野生小蓬竹光合参数及其测量数据†Table 2 The simulated photosynthetic parameters fitted by five models of wild D.luodianense and the measured data
综上所述,采用修正的直角双曲线模型对于拟合小蓬竹光合作用光响应曲线最为适宜。因此,野生小蓬竹各项光合参数采用修正的直角双曲线模型的拟合值。野生小蓬竹光饱和点(1 750.75 μmol·m-2s-1)较高,比同为小型丛生竹种的缺苞箭竹(900 μmol·m-2s-1左右)[18]高出很多,比大型竹种勃氏甜龙竹[19](1 638 μmol·m-2s-1)也略高,也比高大乔木毛白杨(760~1 100 μmol·m-2s-1)[20]和毛红椿(1 535.7μmol·m-2s-1)[21]以及高产果树美国山核桃(1 630 μmol·m-2s-1左右)[22]要高,小蓬竹的这一特性决定了其具有较强的光能利用能力,符合阳生喜光植物的特征。同时,其光补偿点(21.40 μmol·m-2s-1)又较低,表明小蓬竹在较弱光照环境下仍能进行有机物积累,因此具有一定的耐荫能力,且对光照的适应范围较宽。
4 讨 论
小蓬竹在造纸、园林绿化以及喀斯特山地植被恢复等应用中具有一定的开发意义,需要对其光合作用特性更进一步研究,这对于保护濒危植物小蓬竹以及贵州喀斯特山区生态恢复与经济发展都具有重要的实际意义。
目前,对光合作用光响应数据的数学处理,以直角双曲线、非直角双曲线应用最为广泛,虽然它们能很好地拟合植物叶片在没有出现光抑制时的光响应数据,但它们都是没有极值的函数,因此无法求取植物最大净光合速率和饱和光强的解析解[15]。在本文中以及其它应用直角双曲线、非直角双曲线进行光响应拟合的研究[23-26]中,利用非线性最小二乘法进行估算得到的最大净光合速率比实测值高出许多,而利用线性方程拟合植物在低光强部分(≤200 μmol·m-2s-1)的光合作用光响应数据得到的直线斜率求取的饱和光强却远小于实测值。同时,由于没有极值的特性也使得这2个模型无法处理植物出现光抑制而净光合速率下降部分的曲线。指数函数在本文的应用效果与其它类似研究[2,15,27]一样,同样存在最大净光合速率与实测值相差较大、无法拟合小蓬竹光抑制部分光响应数据的问题;而按照前人经验假定最大净光合速率的90%[16]或99%[17]所对应的光强为饱和光强,要么远小于实测值,要么无法求取[2,15],显然,这里的假设具有明显的人为性,求取的饱和光强并不是植物本身具有的。二次多项式虽然可以处理植物光抑制条件下的测量数据,但存在没有光补偿点、暗呼吸速率为负值等问题[24,28-29]。
修正的直角双曲线模型[10-11]克服了其它几个模型[5-7,12-13]拟合小蓬竹和其它植物的光合参数时出现的上述各种问题,它不但能直接求取最大净光合速率、饱和光强等参数的解析解,而且求取的参数以及拟合值与实测值吻合得很好[2,15,19,29],这也是该光响应模型与其它模型的最大区别。因此,建议在需要应用数理模型对植物光响应数据进行拟合时采用修正的直角双曲线进行。
此外,在拟合植物光响应数据时由于模型选择的不同,模型间参数混用的现象比较普遍,缺乏统一的定义,导致参数之间的对比性降低。比如直角双曲线、非直角双曲线以及指数函数模型中的α,对于此参数的解释有的研究者认为是表观量子效率,有的则认为是初始量子效率,也有人认为指数函数模型中的α为低光下光化学量子效率而仅作为迭代拟合时的一个系数。本文中按照叶子飘先生的推导[15]把α作为初始量子效率,得到的α值比按照徐大全先生的方法[14]求出的表观量子效率值略大,因此将模型中的α作为表观量子效率可能会偏大。叶子飘和于强先生建议将植物在光补偿点处的量子效率作为表征植物利用光能的一个指标[2],这样可以避免不同模型拟合表观量子效率不具有可比性以及利用徐大全先生的方法求取时可能忽略了植物净光合速率在低光区域的非线性响应问题[30]。
[1] 谢元贵,刘济明,陈 洪,等.不同喀斯特生境小蓬竹无性系构件研究[J].西南大学学报:自然科学版,2009,31(6):46-50.
[2] 叶子飘,于 强.光合作用光响应模型的比较[J].植物生态学报 ,2008,32(6):1356-1361.
[3] Robert E S, Mark A, John S B.Kok effect and the quantum yield of photosynthesis[J].Plant Physiology,1984,75:95-101.
[4] Blackman F F , M A,D S C.Optima and limiting factors[J].Annals of Botany,1905,os-19(2):281-295.
[5] Baly E C.The kinetics of photosynthesis[J].Proceedings of the Royal Society of London Series B (Biological Sciences),1935,1:218-239.
[6] Thornley J H M.Mathematical Models in Plant Physiology[M].London: Academic Press(Inc.),1976:86-110.
[7] Bassman J, Zwier J C.Gas exchange characteristics of Populus trichocarpa, Populus deltoids and Populus trichocarpa × P.deltoids clone[J].Tree Physiology, 1991,8:145-159.
[8] Prado C H B A, Moraes J A P V.Photosynthetic capacity and specific leaf mass in twenty woody species of Cerrado vegetation under field condition[J].Photosynthetica, 1997,33(1):103-112.
[9] Watling J R, Press M C, Quick W P.Elevated CO2induces biochemical and ultrastructural changes in leaves of the C4 cereal sorghum[J].Plant Physiology, 2000,3:1143-1152.
[10] Ye Z P.A new model for relationship between light irradiance and the rate of photosynthesis in Oryza sativa[J].Photosynthetica,2007,45(4):637-640.
[11] Ye Z P, Yu Q.A coupled model of stomatal conductance and photosynthesis for winter wheat[J].Photosynthetica, 2008,4:637-640.
[12] 郭水良,方 芳,黄 华,等.外来入侵植物北美车前繁殖及光合生理生态学研究[J].植物生态学报,2004,28(6):787-793.
[13] 刘锦春,钟章成,何跃军,等.重庆石灰岩地区十大功劳(Mahonia fortunei)的光合响应研究[J].西南师范大学学报:自然科学版,2005,32(2):158-163.
[14] 许大全.光合作用效率[M].上海:上海科学技术出版社,2002:2-52.
[15] 叶子飘.光合作用对光和CO2响应模型的研究进展[J].植物生态学报 ,2010,34(6):727-740.
[16] 王满莲,冯玉龙,李 新.紫茎泽兰和飞机草的形态和光合特性对磷营养的响应[J].应用生态学报,2006,17(4),602-606.
[17] 黄红英,窦新永,孙蓓育,等.两种不同生态型麻疯树夏季光合特性的比较[J].生态学报,2009,29(6):2861-2867.
[18] 王亚萍,王开运,张远彬,等.缺苞箭竹气体交换对CO2浓度及光照强度的响应[J].西北林学院学报,2008,23(1):6-9.
[19] 李伟成,王树东,钟哲科,等.几种经验模型在丛生竹光响应曲线拟合中的应用[J].竹子研究汇刊,2009,28(3):20-24.
[20] 富丰珍,徐程扬,李广德,等.冠层部位对三倍体毛白杨光合生理特性的影响[J].中南林业科技大学学报,2010,30(3):95-99.
[21] 赵 坤,吴际友,陈 瑞,等.毛红椿光合及水分生理生态特性[J].中南林业科技大学学报,2011,31(5):87-91.
[22] 吕芳德,黄 菁,和红晓.不同树龄美国山核桃光合作用的试验与分析[J].中南林业科技大学学报,2011,31(3):44-46.
[23] Kyei-Boahen S, Lada R, Astatkie T, et al.Photosynthetic response of carrots to varying irradiances[J].Photosynthetica,2003,41(2):301-305.
[24] 高 峻,孟 平,吴 斌,等.杏—丹参林药复合系统中丹参光合和蒸腾特性的研究[J].北京林业大学学报, 2006, 28(2):64- 67.
[25] Leakey A D B, Uribelarrea M, Ainsworth E A, et al.Photosynthesis, productivity,and yield of maize are not affected by Open-Air elevation of CO2concentration in the absence of drought[J].Plant Physiology, 2006,2:779-790.
[26] 王照兰,杨 持,杜建材,等.不同生态型扁蓿豆光合特性和光适应能力[J].生态学杂志,2009,28(6):1035-1040.
[27] Yu Q, Zhang YQ, Liu YF, et al.Simulation of the stomatal conductance of winter wheat in response to light, temperature and CO2changes[J].Annals of Botany,2004,93(4):435-441.
[28] 刘宇锋,萧浪涛,童建华,等.非直线双曲线模型在光合光响应曲线数据分析中的应用[J].中国农学通报,2005,121(8):76-79.
[29] 叶子飘,于 强.一个光合作用光响应新模型与传统模型的比较[J].沈阳农业大学学报,2007,38(6):771-775.
[30] Prioul J L, Chartier P.Partitioning of transfer and carboxylation components of intracellular resistance to photosynthetic CO2fixation: a critical analysis of the methods used[J].Annuals Botany,1977,41(4):789-800.
Model fitting on light response curve of photosynthesis of wild Drepanostachyum luodianense
LIAO Xiao-feng, LIU Ji-ming, ZHANG Dong-kai, JIN Yong, ZHANG Yong, YAN Guo-hua, WANG Min
(Forestry College of Guizhou University, Guiyang 550025, Guizhou, China)
Several typical models of light-response curve of leaf net photosynthesis, such as non-rectangular hyperbolic model,rectangular hyperbolic model, prompted rectangular hyperbolic model and index function model, were tested on wild Drepanostachyum luodianense, and the applicability of these models also were investigated.The results were summarized as follows:① the plant’s analytical solution of maximum net photosynthetic rate (Pnmax) and saturation point (Isat) couldn’t be solved by rectangular hyperbola model, non-rectangular hyperbola model and exponential equation model, while the values simulated by combining other methods existed a big difference compared with the measured value, and the data under photoinhibition couldn’t be processed by these three models; ②the quadratic polynomial model could process the data under photoinhibition to a certain extent, but the photosynthetic parameters simulated by it also existed larger difference compared with the measured value, even generated logic error; ③the photosynthetic parameters simulated by modified rectangular hyperbola model all were close to the measured values ,and this model also could well process the data under photoinhibition; ④the Pnmax, Isat, compensation point(Ic), respiration rate(Rd) and initial quantum efficiency(a) of wild D.luodianense simulated by modified rectangular hyperbola model were 8.53 µmol·m-2s-1, 1 750.75 µmol·m-2s-1,21.40 µmol·m-2s-1, 1.06 µmol·m-2s-1and 0.054 respectively.
Drepanostachyum luodianense; light-response curve; light-response model; modified rectangular hyperbola
2011-08-29
贵州省科技厅对外合作项目“濒危植物小蓬竹生态适应性及育苗技术研究”[黔科合外G字(2009)70010];贵州大学引进人才基金项目“濒危植物小蓬竹耐旱性及小生境生长特征研究”;贵州大学大学生SRT项目“喀斯特濒危植物小蓬竹耐旱性研究”
廖小锋(1987—),男,重庆梁平人,硕士生,从事植物生理生态学研究
刘济明(1963—),男,重庆市人,教授,博士,硕士生导师,主要从事植物生理生态学研究;E-mail: karst0623@163.com
S718.43
A
1673-923X(2012)03-0124-05
[本文编校:谢荣秀]
