湖北枝江市石子岭水库蓝藻水华研究
2012-12-25汪志聪饶本强李印霞刘永定李敦海中国科学院水生生物研究所湖北武汉430072中国科学院研究生院北京00049信阳师范学院生命科学学院河南信阳464000
汪志聪,饶本强,李印霞,刘永定,李敦海* (.中国科学院水生生物研究所,湖北 武汉 430072;2.中国科学院研究生院,北京 00049;3.信阳师范学院生命科学学院,河南 信阳 464000)
湖北枝江市石子岭水库蓝藻水华研究
汪志聪1,2,饶本强1,3,李印霞1,2,刘永定1,李敦海1*(1.中国科学院水生生物研究所,湖北 武汉 430072;2.中国科学院研究生院,北京 100049;3.信阳师范学院生命科学学院,河南 信阳 464000)
2009年8月21日对枝江市作为饮用水源的石子岭水库浮游植物和水体营养盐状况进行了调查.结果表明,该水库已经发生了微囊藻水华;浮游植物群落结构定量分析和多样性指数计算表明石子岭水库明显富营养化,惠氏微囊藻是是群落中绝对优势种.同时浮游植物生态位测度值表明微囊藻等蓝藻在时下的生态因子作用下将呈现逐渐衰退,而绿藻种群扩张,水库中浮游植物明显呈现由蓝藻优势向绿藻优势群落演替趋势.测定叶绿素荧光参数,以了解微囊藻水华的生长状态,比较研究中午强光下和经过24 h暗恢复及与室内培养的惠氏微囊藻三者的差异,表明夏末秋初阶段水库中微囊藻受到了一定的光照胁迫.水库中营养盐分析表明过高的总氮、总磷浓度和适宜的 N/P比可能是水华暴发的主要原因,同时也是导致石子岭水库浮游植物呈现特定的群落结构特征的关键因素.
石子岭水库;蓝藻水华;浮游植物;生态位宽度;生态位重叠
枝江市位于湖北省中部偏西,地处长江中游北岸.东与荆沙接壤,西与宜昌毗邻,属副亚热带,年均气温15.9~16.5℃,日照1500~2000h,无霜期220d左右,年均降雨量1032mm.石子岭水库是枝江市水利局直属的中型水库,距城关26km.水库承用面积49.6km2,库容2.17×107m3,养鱼面积l.0×106m2,灌溉农田53.3km2[1],是一座以农业灌溉为主、兼营养鱼以及村镇生活用水的综合型水库.
石子岭水库一直以来就是当地居民主要饮用水源地,此前一直未发现有蓝藻水华现象,水质状况良好.从2009年7月中旬开始至本调查期间石子岭水库一直出现蓝藻水华,严重影响了问安镇自来水厂对该镇 1.5万居民日常用水供应.但迄今关于作为饮用水源地的水库突然出现严重蓝藻水华的现象还鲜为报道.为了对水库水体污染状况做出正确评估和提出切实有效的水质改善措施,笔者于2009年8月21日首次调查了石子岭水库中浮游植物群落结构和总氮总磷营养盐状况,并以毗邻的熊家港水库水体相应指标作对照说明,拟从该水库微囊藻的群落结构特征和野外条件下的光合特性以及水体氮磷含量等方面探讨石子岭水库蓝藻水华发生的原因和机理,为水华防治提供基本的实验依据.
1 样点设置与分析方法
1.1 采样点设置和实验方法
分别在水库的上游(3#)和下游(1#、2#、4#)各水域设置了4个采样点,并在紧邻的熊家港水库设置了对照点(5#),点位设置如图1所示.

图1 石子岭水库与熊家港水库Fig.1 Shiziling Reservoir and Xiongjiagnag Reservoir
1.2 叶绿素测量
取100mL水样,经1.2μm的GF/C玻璃纤维滤膜过滤后,于4℃保藏,并在24h内带回实验室分析.滤膜经80%的丙酮冰浴充分研磨,后将研磨样品转入10mL的离心管中,定容终体积约8mL.置于4℃下浸提20~24h后,浸提液10000r/min离心10min,上清液用于叶绿素测量.用10mm光径的比色皿装样置入Ultrospec3000紫外可见分光光度计测量上清液在665,649nm 2波段的吸光度值.叶绿素a计算公式参考Vernon的方法[2]Chla= 11.63OD665-2.39OD649,μg/L.
1.3 浮游植物群落结构定量分析
样品采集后,立即用鲁哥试液固定,沉降48h后,弃去上清液,底层沉降藻混匀后,用Leica双筒光学显微镜对藻进行定性定量分析.计算微囊藻群体密度和群体平均所含细胞数时,至少取50个视野,计数至少300个群体含有细胞量的平均值.生态位宽度利用 Levins公式[3]计算,生态位重叠利用 Petraitis公式[4]计算.多样性指数公式为:
Shannon-Wiener多样性指数[5]:
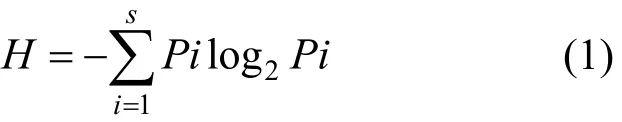
Simpson多样性指数[6]:
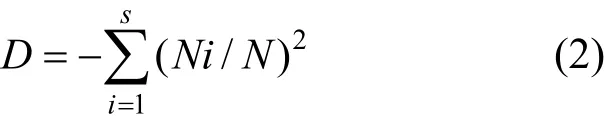
式中:Ni是群落中第i个物种的数目,N是群落中物种总数目,Pi=Ni/N.
1.4 叶绿素荧光参数
用 PAM2100(WALZ, Germany)测定叶绿素荧光参数[7],在光化光的强度(1800μmol/(m2·s))上稍作修改.样品经5min(不宜过长)的暗适应后,在小于 0.15μmol/(m2·s)的测量光(ML)下测定最大叶绿素荧光Fv/Fm,打开光化光在256μmol/(m2·s)使藻进行光适应,做快速光响应曲线,得出ΦPSII、qN和qP的值.在1800μmol/(m2·s)的条件下模拟当时野外实测强光条件下,测量藻类在环境光下的光合系统活性.
1.5 总氮、总磷分析
总氮采用碱性过硫酸钾消解紫外分光光度法测定,总磷用钼酸铵分光光度法测定,详细方法参见文献[8].
1.6 统计分析方法
每个的叶绿素荧光数据每个样品做5个平行测量,取平均值;氮磷营养盐浓度数据做3个平行测量,同样取平均值.SAS统计分析软件平台上分析测量数据.方差分析(One-way ANOVA)检验用来分析在样品之间是否有显著的差异,P <0.05定为显著水平.
2 结果与讨论
2.1 石子岭水库叶绿素含量分析
随着城市经济的发展和乡村城镇化的改革,众多的淡水湖泊呈现明显的富营养化和蓝藻水华现象[9-11].水库作为一种农业灌溉设施,也承担着居民饮用水源的重要作用,但水库的富营养化也是一个目前已成为威胁民生的一个日益突出的问题[12].图2中显示石子岭水库中叶绿素的含量平均为 91.69μg/L,在水库东角的 4#小库湾中叶绿素浓度高达 163.09μg/L,肉眼可见已经出现表面水华现象.
相邻的熊家港水库水体明显为绿色,但透明度很高,肉眼也不可见颗粒状藻,浮游植物网采也收集不到藻颗粒.经检测叶绿素含量高达70.21μg/L(图 2),镜检优势种是绿藻,次优势种是微囊藻.
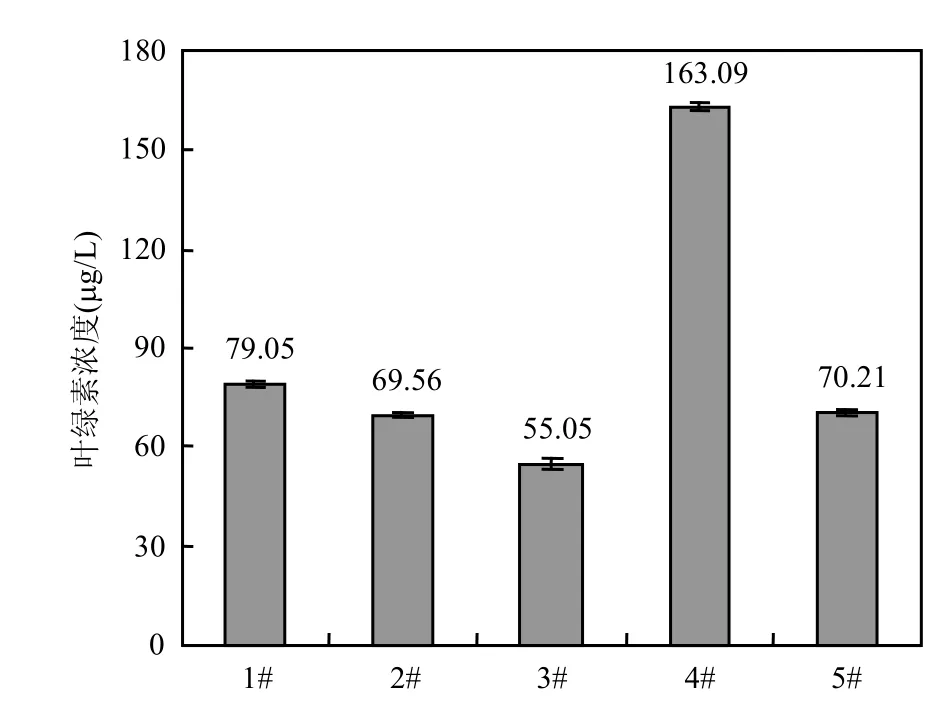
图2 石子岭水库水体叶绿素含量Fig.2 Chlorophyll a concentration of all sampling points in the reservoirs
石子岭水库有明显的蓝藻水华,虽然不像滇池、巢湖和太湖所出现的表面聚集水华,但肉眼可见水体中藻颗粒密度较大,并散发出微囊藻特有的臭味.以叶绿素水平来衡量,石子岭水库已经暴发了轻度蓝藻水华.相邻的熊家港水库水体明显为绿色,虽然不见成群体的藻颗粒,但以叶绿素为衡量标准,根据水华定义的研究[13]确定发生了绿藻和蓝藻共同水华.
2.2 石子岭水库浮游植物群落结构定量调查
样品经固定后于实验室光学显微镜下进行浮游植物鉴定,结果见表 1,水库中微囊藻在群落中占绝对优势,约占总生物量的98%以上,而在作为对照的熊家港水库(5#)中却以绿藻门的螺旋纤维藻占优势,微囊藻在数量上仅占21%.
Shannon-Wiener和Simpson 2种多样性指数均显示(表2)石子岭水库的多样性指数较低,均在1.638以下,最低达到0.012.上游3#的多样性指数略高于下游的4#,但显著低于下游的1#、2#,并且石子岭水库的多样性指数显著地低于熊家港水库,水库上游 3#的群落多样性比下游稍高一些,微囊藻的密度也更大(表1).5#熊家港水库发生了微囊藻和纤维藻共同占优势的水华,库中浮游植物多样性较石子岭水库显著的高.石子岭水库的生物多样性指数较低,微囊藻这种富营养化物种是群落中绝对优势种.这表明水库水质已经出现一定程度的富营养化.水库上游群落多样性指数比下游位点更高,表明水库上游的水质富营养化更严重,水体营养物质释放源可能在入库的主河道区.熊家港水库发生了蓝、绿藻水华,群落结构特征与石子岭水库有明显的区别,生物多样性显著的高于石子岭水库.
表 3显示了石子岭水库浮游植物优势种的生态位宽度和生态位重叠值,表明惠氏微囊藻(Microcystis wesenbergii)、 细 小 平 裂 藻(Merismopedia minima)、小席藻(Phormidium tenue)等 蓝 藻 和 四 尾 栅 藻 Scenedesmus quadricauda等绿藻的生态位较宽,而生态位重叠值 均 为 负 值 .单 角 盘 星 藻 (Scenedesmus quadricauda)、空球藻(Eudorina elegans)、湖生小椿藻(characium limneticum)等绿藻虽然生态位宽度较小,但生态位重叠值为正值.虽然四尾栅藻Scenedesmus quadricauda生态位宽度较宽,但是生态位重叠呈负值.石子岭水库中蓝藻的生态位较宽,在水体中分布较普遍,适应性较强,而生态位重叠值均为负值,表明蓝藻虽然适应性较强,但生态因子的演变将不利于蓝藻物种的进一步扩张,微囊藻和平裂藻在后期将会呈现衰退的趋势.绿藻虽然生态位宽度较小,但生态位重叠测度值较高.这表明绿藻虽然分布区域不如蓝藻物种广泛,但在后期生物量将会增加;蓝绿藻的更替正在进行.各个局部位点的微囊藻水华的生物量与群体形态大小呈正相关的(图3),这表明较大的群体,浮力更大,更容易在湖泊某些区域聚集,形成肉眼可见的水华现象.
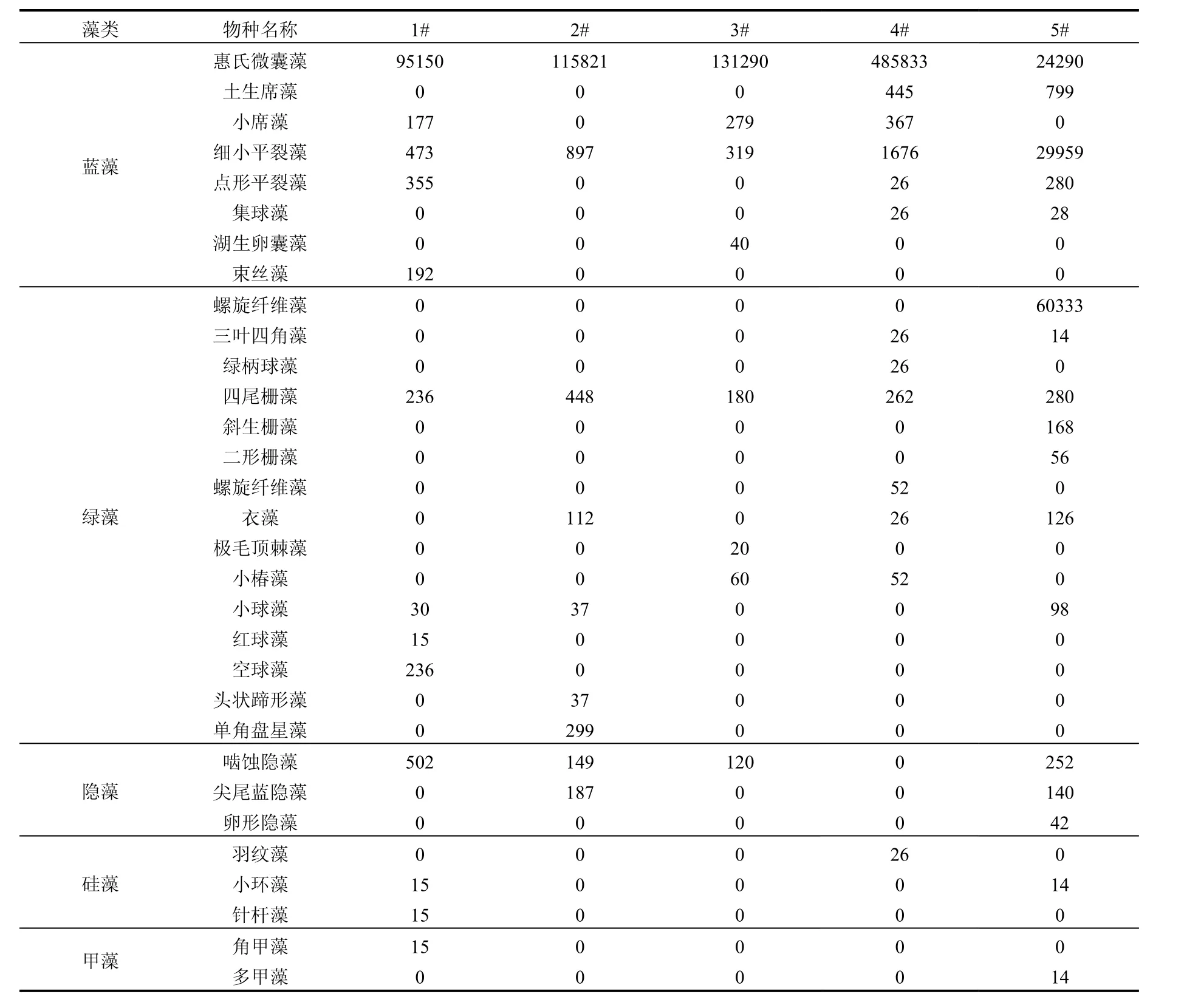
表1 石子岭水库浮游植物群落结构(×103个细胞/L)Table 1 Phytoplankton community structure of Shiziling Reservoir (×103 cells/L)
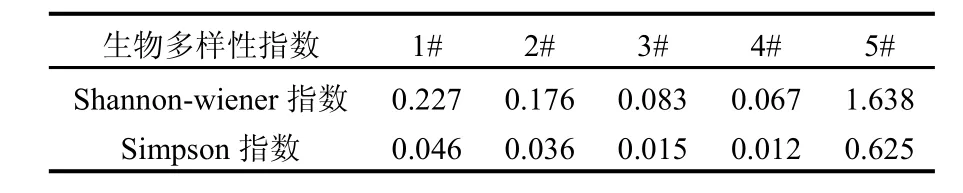
表2 石子岭水库浮游植物群落Shannon-Wiener和Simpson多样性指数Table 2 Shannon-Wiener and Simpson’s diversity indexes of phytoplankton community in Shiziling Reservoir
2.3 石子岭水库中微囊藻群体密度和细胞含量
显微镜下观察,微囊藻为单一种,全部为惠氏微囊藻,在可见表面水华的石子岭水库东角4#采样点,计数后得微囊藻的群体为 3.7×103个/mL,远高于水库其他未见藻体上浮的水域,同时 4#样点的每个群体平均细胞数目也较其它水域显著大(图3).但与铜绿微囊藻占优势的太湖群体(数据待发表)相比,每个群体平均细胞数上要小得多.出现表面水华的 4#样点微囊藻的群体较多,群体含有的细胞数较其他样点的显著 要高.

表3 石子岭水库浮游植物优势种的生态位宽度和生态位重叠Table 3 Niche breadth and niche overlap of dominant phytoplankton species in Shiziling Reservoir
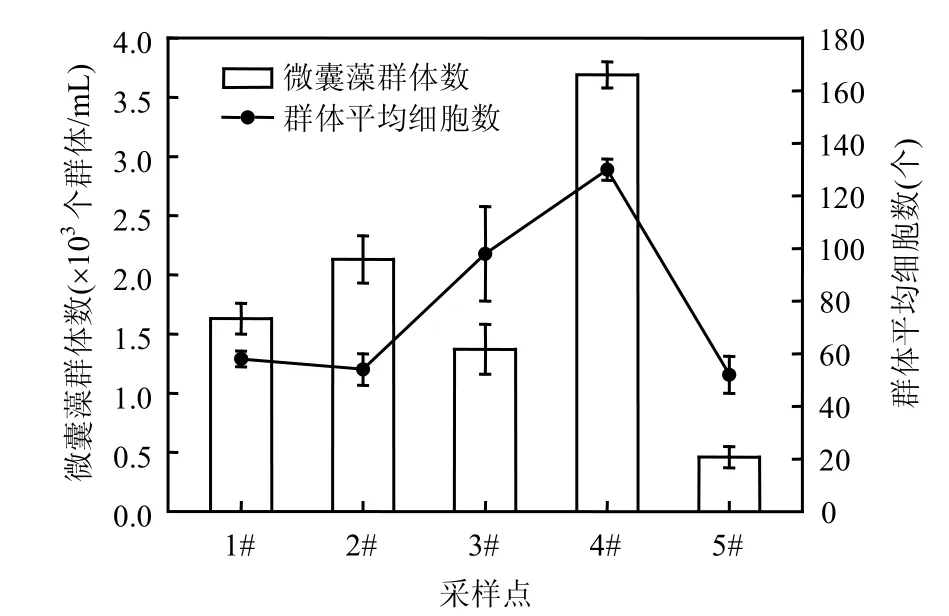
图3 石子岭水库中微囊藻群体分析Fig.3 The analysis of Microcystis colony in Shiziling Reservoir
2.4 叶绿素荧光参数分析
由图4可见,温室内培养的微囊藻是在各种条件适宜的情况下生长的,光合系统的光合活性相对较高.最大光化学效率(Fv/Fm)、实际光化学效率(ΦPSII)和光化学淬灭(qP)分别达到 0.446± 0.008、0.467±0.010、0.856±0.048,而非光化学淬灭(qN)较小.野外湖泊中微囊藻在模拟现场高强度光化光条件下,光合活性显著下降(one-way ANOVA, P<0.05),Fv/Fm、ΦPSII和 qP分别为0.259±0.006,0.089±0.021和 0.383±0.053,而反应中心过剩激发能qN显著升高到0.460± 0.076.现场采集的微囊藻经过48h的暗适应恢复后,非光化学qN消失,光合活性Fv/Fm、ΦPSII和qP有所升高但依然处于一个相对较低的水平,显著低于室内同种微囊藻的光合活性.
石子岭水库各点的藻均显黄绿色,浮游植物网采悬浮的黄绿色藻颗粒清晰可见.最大光化学量子产量 Fv/Fm是一个很稳定的值,藻类的比高等植物的低,约为 0.65[14].当藻类受到环境胁迫时,Fv/Fm显著下降[15],因此 Fv/Fm是研究光抑制或各种环境胁迫对光合作用影响的重要指标.
图4的叶绿素荧光参数中,作为对照的室内培养的惠氏微囊藻的Fv/Fm为0.446,Tytler等[16]也报道的适宜情况下微囊藻荧光参数范围为0.41~0.47,然而野外微囊藻在中午较强光照条件下Fv/Fm为0.256,表明在光合活性有所下降,在一定程度上受到了光胁迫.qP表示开放的 PSⅡ反应中心所占的比例[17],ΦPSⅡ表示PSⅡ光化学能量转化的有效量子产量[18],qN表示藻对过剩光能的耗散能力[19].在未受到环境胁迫条件下,qN较低,几乎接近于0,ΦPSII和qP较高.野外微囊藻实时荧光比实验室内适宜条件下培养的同种惠氏微囊藻相应的荧光参数显著下调,qN为0.460,显著高于适宜条件下的 qN(0.002).在适宜条件下,qP较大,qN较小,当藻受到环境胁迫时,qP下降,qN上升,当环境胁迫强度过大、时间过长,藻甚至会丧失光合活性,相应的 qP和 qN都变小,甚至降低为 0,此时藻样即使经充分暗适应后,光合活性也无法恢复.
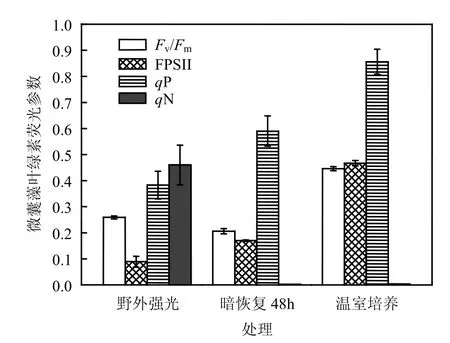
图4 石子岭水库微囊藻叶绿素荧光参数Fig.4 Chlorophyll fluorescence parameters of Microcystis in Shiziling Reservoir
现将野外的微囊藻样品经室内 25℃、黑暗条件下恢复48h后,其Fv/Fm进一步降低为0.206,但ΦPSII较暗恢复前有所上升,光化学淬灭qP也增大了,非光化学 qN下降至接近于 0.这是因为蓝藻光合作用分类为藻胆体(PBS)生物, PQ是光合作用和呼吸作用的共同电子传递体,藻在长时间的暗适应下光合作用向状态 2转换,表现为Fv/Fm略有下降[20],而在暗适应中,由于叶绿素荧光的暗弛豫过程,非光化学淬灭 qN下降,氧化态的 QA黑暗中逐渐被还原,光合电子传递链抑制得以恢复,从而使光化学淬灭 qP提高.上述暗恢复的结果进一步表明水库中微囊藻受到了环境因子的胁迫作用.推测可能是水库透明度较高,过强的光线及表层水温的上升使藻的生长受到一定的抑制,并且这种抑制可以导致藻的光合活性不可逆的失活,并在秋后季节逐步衰退.值得庆幸的是发生的是非产毒特性的惠氏微囊藻株系,如果是产毒株系水华,那这种水华及衰退后的细胞溶解所释放出来的毒素对水体生态系统和人类健康都是一个较大的威胁.因此,我们不仅要关注石子岭水库中富营养化进程,争取尽量控制营养物污染染头,还需要实时监测这个已经富营养化水库中水华蓝藻群落结构的特别是产毒蓝藻株系的变化.
2.5 石子岭水库总氮和总磷含量分析
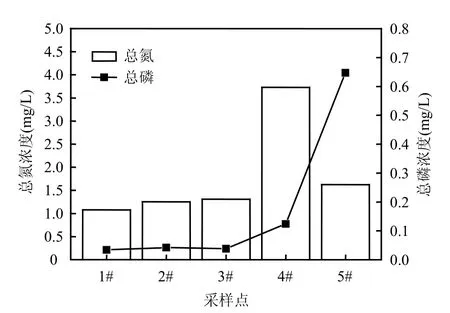
图5 石子岭水库总氮与总磷浓度Fig.5 The concentration analysis of total nitrogen and total phosphorus in the two reservoirs
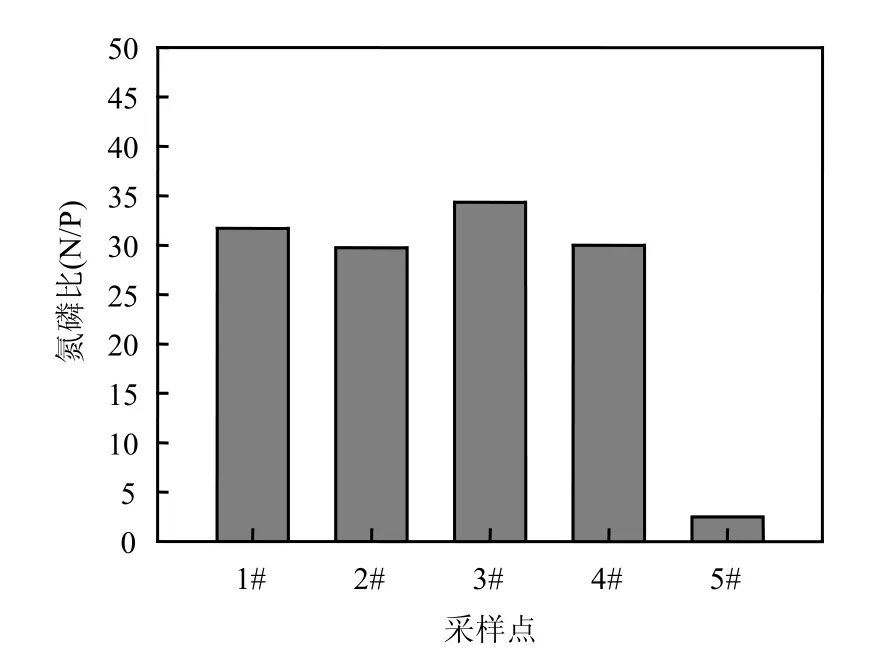
图6 石子岭水库氮磷比值的比较Fig.6 The ratio of nitrogen to phosphorus among all the sampling points
前面两水库中群落结构差异与它们的富营养状况是紧密相关的.作为饮用水源方面,石子岭水库氮磷已经严重超标,依据GB3838-2002水质分类标准,石子岭水库总氮平均为1.84mg/L,达到Ⅴ类水质标准,总磷为0.06mg/L,达到Ⅳ水质标准(图 5).石子岭水库东角 4#点总磷浓度高达0.124mg/L,表明水库部分水域总磷达到达到Ⅴ水质标准.而对照样点5#熊家港水库中富营养化更严重,总磷为0.647mg/L,显著高于石子岭水库,约为其平均水平的10.8倍.石子岭水库各样点的N/P均在30左右,而熊家港水库因为总磷浓度非常高,总氮浓度偏低,导致N/P比值降低至2.51(图6).这种水体中营养状态是决定浮游植物群落结构组成的重要因素之一[21],其中氮磷比对水华蓝藻优势形成有很大的影响[22].在紧密相邻的石子岭水库和熊家港水库中,地理环境和气候等众多的生态因子是相同的,这种营养盐浓度和组成的差异可能是造成两水库中浮游植物群落结构显著不同的主要原因.
3 结论
3.1 依据叶绿素含量和浮游植物生物量两种水华界定标准,石子岭水库中叶绿素含量达91.69μg /L,确定已经暴发了微囊藻水华,而在地理位置上毗邻的熊家港水库暴发微囊藻和纤维藻共同优势的水华.
3.2 藻类物种数目较少、富营养化物种微囊藻是绝对优势种和生物多样性指数较低(最小为0.012),表明石子岭水库各库区均呈现明显的富营养化现象.群落生态位测度值和叶绿素荧光参数值较低(0.256),均表明水库中浮游植物群落结构呈现蓝藻优势向绿藻优势的演替趋势.
3.3 石子岭水库中总氮(平均为 1.84mg/L)、总磷(平均为 0.06mg/L)营养盐浓度分析表明水库水质污染严重,多为Ⅳ、Ⅴ类水质,熊家港水库营养盐污染更严重.营养盐组成差异可能是两水库浮游植物群落结构不同的重要原因.
[1] 梁开学,梁明德.枝江县石子岭水库渔业安置渔民规划报告 [J].水利渔业, 1995,2:39-42.
[2] Vernon L P, Seely G R. The Chlorophylls [M]. New york, 1966.
[3] Levins R. Evolution in changing environments: some theoretical explorations [M]. USA: Princeton University Press, 1968.
[4] Petraitis P S. Likelihood measures of niche breadth and overlap [M]. Ecology, 1979,60:703-710.
[5] Shannon E E, Weaver W. The mathematical theory of communication [M]. Urbana, Chicago, London: University Illinois Press, 1949:125.
[6] Simpson E H. Measurement of diversity [J]. Nature, 1949,163: 688.
[7] Wang Z C, Zuo M, Wang Y C, et al. Dynamics of chlorophyll fluorescence and eco-morphological properties of Microcystis bloom in Meiliang Bay of Lake Taihu, China [J]. Fresenius Environmental Bulletin, 2011,20(9):2295-2305.
[8] Wang Y C, Wang Z C, Wu W J, et al. Seasonal regime shift of an alternative-st ate Lake Xingyun, China [J]. Fresenius Environmental Bulletin, 2010,19(8):1474-1485.
[9] Yang H, Xie P, Xu J, et al. Seasonal variation of microcystin concentration in Lake Chaohu, a shallow subtropical lake in the People's Republic of China [J]. Bulletin of Environmental Contamination and Toxicology, 2006,77:367-374.
[10] 刘聚涛,杨永生,姜加虎.太湖蓝藻水华灾害风险分区评估方法研究 [J]. 中国环境科学, 2011,31(3):498-503.
[11] Zhang M, Xu J, Xie P. Nitrogen dynamics in large shallow eutrophic Lake Chaohu, China. [J]. Environmental Geology, 2008, 55:1-8.
[12] 许秋瑾,郑丙辉,朱延忠,等.三峡水库支流营养状态评价方法[J]. 中国环境科学, 2010,30(4):453-457.
[13] 郑建军,钟成华,邓春光.论水华的定义 [J]. 水资源保护, 2006, 2(5):45-47.
[14] kolber Z, Zehr J, Falkowski P G. Effects of growth irradiance and nitrogen limitation on photosynthesis energy conversion in photosystem Ⅱ[J]. Plant Phsiol., 1988,88:72-79.
[15] 许大全, 张玉忠, 张荣铣. 植物光合作用的光抑制 [J]. 植物生理学通讯, 1992,28(4):237-243.
[16] Tytler E M, Whitelam G C, Hipkins M F, et al. Photoinactivation of photosystem II during photoinhibition in the cyanobacterium [J]. Planta, 1984,60(3):229-234.
[17] Baker N R. Chlorophyll fluorescence: A probe of photosynthesis in vivo [M]. Annu. Rev. Plant Biol., 2008,59:89-113.
[18] Maxwell K, Johnson G N. Chlorophyll fluorescence—a practical guide [J]. Journal of Experimental Botany, 2000,51(345):659-668.
[19] 冯志立,冯玉龙,曹坤芳.光强对砂仁叶片光合作用光抑制及热耗散的影响 [J]. 植物生态学报, 2002,26(1):77-82.
[20] 杨胜铭,高辉远,邹 琦.状态转换对光合作用中激发能分配的调节及其与光破坏防御的关系 [J]. 植物生理学报, 2001,37(2): 89-94.
[21] Lalli C M, Parsons T R. Biological oceanography: An Introduction [M]. New York: Pergamon Press, 1993:45-79.
[22] 许 海,朱广伟,秦伯强,等.氮磷比对水华蓝藻优势形成的影响[J]. 中国环境科学, 2011,31(10):1676-1683.
Primary study on cyanobacteria bloom in Shiziling Reservoir of Zhijiang, Hubei Province.
WANG Zhi-cong1,2, RAO Ben-qiang1,3, LI Yin-xia1,2, LIU Yong-ding1, LI Dun-hai1*(1.State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China; 2.Graduate University of Chinese Academy of Sciences, Beijing 100049, China;3.College of Life Sciences, Xinyang Normal University, Xinyang 464000, China). China Environmental Science, 2012,32(10):1875~1881
A survey of phytoplankton and trophic status was carried out on August 21, 2009 in Shiziling Reservoir of Zhijiang which was used as a source of drinking water. The results showed that the reservoir had occurred Microcystis bloom. The quantitative analysis and diversity indices of phytoplankton community showed that the whole reservoir was in obvious eutrophic state, and Microcystis wesenbergii (Komárek) Komárek was the absolute dominant species. The niche measurement of phytoplankton indicated that cyanobacteria including Microcystis were being declining while Chlorophyta populations was being expanding, and the community successional tendency between Cyanobacteria and Chlorophyta was obvious. To understand the growth state, the chlorophyll fluorescence parameters of Microcystis sampled from the reservoir, cultured in dark for 24h or cultured underlaboratory illuminated conditions were compared. The results showed that in the late summer and early autumn Microcystis in Shiziling reservoir suffered from strong light stress. The analysis of nutrients suggested that the high concentrations of total nitrogen and total phosphorus and the suitable value of N/ P ratio were the main reasons for the occurrence of Microcystis bloom, and also were the key factors that resulted in the specific characteristics of phytoplankton community characteristics in Shiziling Reservoirs.
Shiziling Reservoir;cyanobacteria bloom;phytoplankton;niche breadth;niche overlap
2011-12-24
国家“973”项目(2008CB418002)和河南省高校青年骨干教师资助计划项目(2010GGJS-106)联合资助.
* 责任作者, 李敦海, lidh@ihb.ac.cn
X832
A
1000-6923(2012)10-1875-07
汪志聪(1984-),男,湖北鄂州人,中国科学院水生生物研究所博士研究生,主要从事藻类环境生物学和水体生态学研究.发表论文10余篇.
