鲕粒成因研究的新进展①
2012-12-14梅冥相
梅冥相
(中国地质大学地球科学与资源学院 北京 100083)
鲕粒成因研究的新进展①
梅冥相
(中国地质大学地球科学与资源学院 北京 100083)
鲕粒的成因一直是一个谜一样的沉积学难题.rehm等在2006年的实验室研究的结果表明,将鲕粒的形成可以与叠层石进行类比,是一个特殊的球状微生物席的产物,从而将鲕粒归为微生物成因。最近,来自于巴哈马现代鲕粒的研究,Duguid等在2010年认为,鲕粒形成与微生物活动不存在一个直接的关系,重新强调了鲕粒形成的化学过程,即:当鲕粒处在沉积物-水界面(活跃相)的时候,凭借一个在鲕粒上的非晶质钙碳酸盐(ACC)沉淀物的微薄层,鲕粒开始形成;这个ACC微薄层后来结晶成文石针,从而形成新的鲕粒皮层(静止相)。两种观点的直接对立,代表了对鲕粒成因研究的新进展。来自于华南下三叠统的巨鲕、以及华北寒武系鲕粒,特殊的显微组构似乎更多地支持了鲕粒形成的微生物作用机制的学说。
微生物作用 ACC沉淀的化学过程 鲕粒成因 研究进展
0 引言
自从Sorby在1879年认为鲕粒生长就像“滚雪球”一样的认识以来,关于鲕粒形成的理论已经经历了两个世纪的演变。在鲕粒表面导致文石沉淀作用的过程还了解得很少,一般认为,鲕粒的形成需要满足以下条件[1]:1)核心的存在;2)有序更新、并经常脱二氧化碳的相对于钙碳酸盐超饱和的海水;3)一个允许二氧化碳脱气的动荡的环境[2]。虽然曾经推断到,鲕粒皮层形成于海水中的直接沉淀作用、以及生物调节的作用机制[3~5];但是,鲕粒皮层沉淀物的精确形成过程一直处于未知状态。所以说,鲕粒的成因过程还没有得到充分的了解,尽管一些形成过程曾经被总结出[6,7],包括:1)生物过程;2)化学沉淀作用过程;3)物理作用过程;以及4)这些过程可能的综合作用。
鲕粒,常常以平滑的圈层和非有机成因特征如藻丝体等的缺乏而与另一种包覆颗粒——核形石较为容易区分,鲕粒圈层的平滑是区分这两种包覆颗粒的可靠标准,因为在很多鲕粒中也发育显示出有机成因的粘液膜[8]。从Sorby在1879年对鲕粒形成的物理过程描述,到Tucker和Wright[9]的结论——“将鲕粒完全归为物理成因似乎为时过早”,均表明了鲕粒成因所意味着的沉积学难题.rehm等[10,11]花了4年时间,在实验室培养出一个被他们定义的“球状微生物群落”,并人工合成了鲕粒,从而将鲕粒的形成归因为类似与叠层石形成的微生物作用过程。鲕粒形成的微生物成因的假说,又受到最近Duguid等[1]对现代巴哈马鲕粒的观察与研究的挑战,他们认为:鲕粒形成与微生物活动不存在一个直接的关系,重新强调了鲕粒形成的化学过程。两种观点的直接对立,代表了对鲕粒成因研究的新进展。但是,对于形成鲕粒皮层极为重要的非晶质钙碳酸盐(Amorphous calcium carbonate;ACC)微薄层,在Duguid等[1]的鲕粒形成的化学作用机制的学说中没有得到较好的阐释,从而留下了一个令人遗憾的缺陷。来自于华南的下三叠统巨鲕[12],以及华北寒武系滩相浮游鲕粒和叠层石中的底栖鲕粒,似乎更多地支持了Brehm等[10,11]的鲕粒形成的微生物成因假说。因此,对鲕粒成因的研究现状的追索,将为今后的进一步研究提供重要线索而具有较为重要的意义。
1 鲕粒形成的微生物作用假说
与纹层状的叠层石[13]相反,鲕粒具有以下特征: 1)内部通常有一个核心,鲕粒核心可以是生物壳、石英颗粒或其他的较小的碎片(包括一个方解石/文石的聚合作用);2)鲕粒核心由一个方解石或文石晶体的皮层所包围,这些方解石和文石晶体排列成放射状的、切线状的或紊乱状的,而且这些晶体常常排列成同心状纹层;3)鲕粒通常形成在温暖的浅海,水流来回冲洗它们,随着它们的大小的改变而促使它们保持为球形。
在对北海现代微生物席研究[14]的过程中,Brehm等[10]多次观察到了不寻常的活着的生物微球,这些生物微球由丝状蓝细菌(Phormidium sp。)、以及相关联的以一个特定方式排列的硅藻如Navicula属和异养细菌所构成。他们在这些协同的培养基中发现了与化石鲕粒相似形式的碳酸盐沉淀作用,从而提出了一个鲕粒形成的微生物过程的认识和假说[11]。
Brehm等[10,11]报道了在实验室条件下、经历了4年时间所培养出的一个球状微生物群落,以及发生在这个球状微生物群落中的碳酸盐沉淀作用,这个形成鲕粒的碳酸盐沉淀作用过程大致可以总结为以下几个方面:
第一、这种微生物群落代表了一个蓝细菌、硅藻和异养细菌的共生体。
第二、这种特殊的球状微生物群落将形成一个特别的生物微球,球的形成和结构化总是遵循着相同的规则,它随着一个包络层的出现而开始形成,这个包络层包覆着异养细菌和硅藻;紧接着,丝状蓝细菌开始靠近,穿入并在球的内部面排列,这个生物微球可以稳定存在几个月。
第三、在生物微球中,钙化作用结构总是反映了微生物聚集体的形状,这个微生物聚集体产生于反应链中的聚结颗粒的形成作用;也就是说,单个碳酸盐晶体的位置遵循丝状蓝细菌的球形排列而且产生一个空心的球,球状生物膜的结构确定了碳酸盐晶体层的形式:微小的方解石针状体,由呈同心状排列的球形微生物群落所沉积,并且在实验室的培养基中可以连续观察到;这种空心的球可能代表了鲕粒的前体,这就是在纹层状微生物席群落内、在没有硬的核心的情况下所产生的鲕粒的实验室体系。
第四、在生物微球内,晶体形成与微生物生长是平行的,进一步的钙化作用步骤导致了一个稳定的、三维钙化球的出现,在发育完好的集团形成之后上述过程总是反复发生,这个钙化球就是可以与那些在岩石记录之中所见到的鲕粒类比的球状结构。
第五、产生生物微球的微生物群落在实验室已经培养了四年,这些微球的直径可以达到100μm,钙化生物微球的结构清晰地展示出与蓝细菌的鞘相似的同心纹层。原生的球状生物膜的结构确定了球状碳酸盐纹层的形式,在钙化的生物微球中,方解石晶体的组织和排列反映了这些生物微球的原生球状属性。
第六、这些球状结构的聚合体就是著名的鲕粒,代表了先成生物微球的单个鲕粒,将会转变成一个多层的球状碳酸盐聚合体。在培养的晚期阶段,蓝细菌出现在多层的圆形集合体之中,其中沉淀了几个碳酸盐环状物,这种外观与化石鲕粒的薄片图像是可以对比的,而且是钙化生物微球和鲕粒的普遍成因的可视化指征;从而进一步推断到:这些球形结构可能被多次冲洗并且在鲕粒滩中大量聚集,尔后形成鲕粒岩石[16]。
考虑到叠层石是有名的、形成于垂向排列的微生物膜或微生物席的纹层状岩石[13,15],Brehm等[10,11]还进一步论述道,球状微生物席群落肯定表现出一些优势:1)抵抗牧食过程所应该具有的较好的保护作用;2)在水柱中通过球的搬运而得到进一步改进的繁殖作用过程;3)光、矿物和其它资源的较好的组织和利用。这些优势的结果,具有碳酸盐轮圈的一个球状聚合体形成在微生物席内,这些碳酸盐轮圈由许多球形定向的小型方解石晶体所组成,这就可以看作是一个底栖鲕粒.rehm等[10,11]关于实验室培养的以及野外采集的微生物席群落的调查,表明了在正常的垂向纹层状生物膜和生物席上由球状微生物群落造成的鲕粒的形成过程。大多数近代的以及化石的鲕粒是围绕一个核心而形成(球粒、石英颗粒和生物碎屑),但是,在他们的实验中,固体核心对于生物微球的形成以及鲕粒的产生不是必要条件。
在地球化学的钙循环中,微生物的参与是主导地球上中性pH的重要因素。首先出现的碳酸盐沉积形成在前寒武纪,在那个时候微生物主导了生物圈[14,15];在显生宙,通过骨骼生物的骨骼化作用代表的生物矿化作用、以及由此而产生的钙循环成为一个重要的作用过程[17]。在原核生物之中,是有少数的代表如蓝细菌,显示出一个特殊的产生钙碳酸盐沉淀作用的能力[18];进一步讲,钙化作用是由一些蓝细菌类群所特别表现出的一个特性,这些蓝细菌类群如Scytonema、Calothrix、Rivularia、Phormidium、Lyngbya和Plectonema[18],而其他的蓝细菌即使出现在相同水平的碳酸盐饱和状态下也不能产生钙化作用,在结晶作用中,钙化作用所产生的碳酸盐沉淀物经常表现出特殊的属和种的差异;因此,碳酸盐沉淀作用只能发生在一些特定微生物席类型和特定背景之中。
已经认识到,下列生物化学过程会影响碳酸盐饱和状态因而造成碳酸盐沉淀作用,主要包括:1)在光合作用期间由于自养生物(以及可能的化学石化生物)造成的环境碳的消耗;2)在细菌的蛋白质分解过程中的氨基酸的脱氨作用;3)厌氧细菌的异化硫酸盐还原反应;4)硫酸盐还原反应、厌氧脱氮作用、尿素水解作用和甲烷生成作用。因为这些作用将会造成微环境的碱性化作用,但是,对于碳酸盐的形成碱性化作用不是唯一的原因.ammes等[19]曾经提出过特定细菌的特殊属性将产生和影响碳酸盐形成; Contos等[20]认为,在生物底层的带负电荷的点上,不均一的成核作用会促进碳酸盐的沉淀作用,因为这些功能基团能够吸收阳离子如二价钙离子,生物底层是指包括气泡和糖蛋白的脱脂二重层以及蛋白多糖;也就是说,组成生物底层的这些高分子是细菌细胞膜和生物膜(多糖类)的组成物,而且作为钙碳酸盐的成核作用模板[20,21]。这些研究的结果和认识,为Brehm等[10,11]提出的“球状微生物群落形成鲕粒”的假说提供了重要的理论支撑,因为鲕粒可以理解为钙碳酸盐围绕一个核心颗粒的、而且连续不断的同心层聚集作用的结果。
2 鲕粒形成的新的化学作用假说
尽管若干研究的结果提供了鲕粒生物成因引人注目的证据,包括:1)在有机质存在的情况下鲕粒包覆层的人工合成[2,11];2)天门冬氨酸,一种在钙化生物中经常发现的氨基酸,出现在鲕粒之中并且意味着一个活跃的、形成文石的生物过程[22,23];3)再者,包括纳米球的纳米级别的结构已经在鲕粒中识别出而且被解释为细菌成因[4,24,25]。基于对巴哈马现代鲕粒的研究,Duguid等[1]发现,鲕粒形成与微生物活动不存在一个直接的关系,从而重新强调了鲕粒形成的化学过程。这主要来源于以下事实:
第一、鲕粒均具有相同的微生物钻孔交互发育的形式。鲕粒的表面和外部皮层常常被未填充的微生物钻孔过程所间断,然而内部皮层包含有填充微生物钻孔的文石胶结物的两种形态;胶结物形式的两种形态与不同的蓝细菌物种存在关联,一种是 Solentia sp。,另一种被解释为Hyella sp。。
第二、在整个巴哈马,鲕粒的化学特征具有惊人的相似性。所有样品的δ18O和δ13C值直接变化,具有一个近于1‰的变化范围。外部皮层具有较低的δ18O和δ13C值,分别为23.4‰和0.2‰,然而内部皮层则具有较高的值,分别为1.9‰和6.8‰;原因是:在鲕粒内部皮层中,微生物钻孔中的文石胶结物的存在增加了鲕粒的氧和碳同位素总体组成,造成这种现象的原因似乎是与海水接近平衡。再者,在鲕粒皮层内δ18O和δ13C值的同位素变化可能被特征化为一个混合线,这个混合线位于未受到改造的鲕粒皮层以及微生物钻孔的文石胶结物的低值之间。
第三、大多数鲕粒的外部皮层具有较高变化的Mg/Ca比值,而且被解释为一个非晶质钙碳酸盐(ACC)包覆层。外部皮层要比内部皮层具有较高的Mg/Ca比值和较低的Sr/Ca比值,化学上的这个差异是因为在内部皮层之中存在文石胶结物造成的结果。
第四、与电子显微镜(SEM)调查相匹配的稳定同位素和痕迹元素分析的结果表明,微生物在鲕粒的形成之中没有起到关键作用,反而是在鲕粒形成之后改变了鲕粒的化学构成和结构。因此,一个新的鲕粒形成的模式被提出,即当鲕粒处在沉积物-水界面(活跃阶段或活跃相)的时候,鲕粒凭借非晶质钙碳酸盐(ACC)沉淀物的微薄层而开始形成;这个ACC微薄层随后结晶成文石针,而且进一步形成新的鲕粒皮层(静止阶段或静止相)。
基于前人的研究,Duguid等[1]进一步论述道,鲕粒外部皮层中文石的δ18O和δ13C值与海水是不平衡的,这也可能归因于文石的快速沉淀作用[26],或者记录了一个文石针的ACC成因.CC的生物重结晶作用常常发生在很多生物之中,包括棘皮类、双壳类和甲壳类生物之中[27],而且ACC能够重结晶成白云石和文石[28,29],ACC还能够在具有一个非有机的种子晶体的光学连续性中重结晶成文石[29]。因此,有可能文石针在一个先期存在的底层之中发生成核作用,因为它们从一个ACC前体相之中重结晶。根据上述推断和假设,Duguid等[1]进一步推断和描述了鲕粒形成的化学过程:1)鲕粒皮层可能开始是以一个ACC薄层,随后重结晶成文石;2)鲕粒形成过程可以描述为两个阶段:活跃和静止,也就是说,鲕粒皮层的初始沉淀作用(可能以ACC的形式开始)发生在活跃阶段,皮层的进一步形成则发生在静止阶段,在此时此地ACC薄层可能重结晶成文石针;3)一直伴随着鲕粒皮层形成过程的是石内蓝细菌活动,石内蓝细菌在活跃和静止两个阶段均是很活跃的,通过微生物钻孔这些石内蓝细菌正在改变着鲕粒的结构和化学构成,只有当鲕粒被埋藏之后微生物钻孔及其相关的胶结物沉淀作用才会停止,而且蓝细菌再也不能进行光合作用。
鲕粒形成的这个化学作用过程模式,Duguid等[1]认为也可以用来解释放射鲕粒和不规则鲕粒的形成:1)在颗粒碰撞较为普遍的地方,即在一个动荡的环境之中,放射状定向的文石针的发育似乎是不可能的,因为生长的文石针常常被打断而变得不完整[30],在静止阶段ACC的重结晶作用允许细微的放射状晶体的生长,所以在低能的环境中形成放射状鲕粒;2)在弱动荡的环境中,不规则鲕粒的形成也可以运用这个模式来加以解释,对于鲕粒皮层的生长所需要的条件是没有经历悬浮作用,当鲕粒沿着海底运动的活跃阶段就会发育ACC,如果动荡程度较低那么纹层就不会是平的,从而导致了不规则鲕粒皮层的发育。
3 形成鲕粒皮层的ACC的成因
尽管Duguid等[1]的鲕粒形成的新的化学过程假说得到了地球化学和相应的SEM调查的支持,但是,鲕粒形成的这个新的化学作用模式的基本前提是: “鲕粒皮层可能开始是一个ACC薄层,随后重结晶成文石”.uguid等[1]没有涉及到非晶质钙碳酸盐(ACC)的来源,所以,他们提出的鲕粒形成的化学作用模式留下了一个最基本的理论缺陷。
最为关键的问题是ACC本身是如何形成的.ddadi等[27]论述道:1)纯净的非晶质钙碳酸盐(ACC)是高度不稳定的,然而一些生物也会产生稳定的ACC,所知道的原因是ACC具有作为更稳定的晶质文石和方解石的过渡性前体的功能;2)生物成因的ACC的研究表明,ACC存在较为明显的结构差异,其稳定形式是含水的,而过渡形式则不含水;3) ACC能够形成在生物体外以很多不同的形式形成,为涉及到ACC转变成钙碳酸盐的结晶形式的稳定化作用、去稳定化作用和转变作用的可能机制带来了一线曙光;4)已经知道的事实是,幼虫阶段的棘皮动物和软体动物确实使用了ACC作为前体相来进行生物矿化(骨骼化)作用,而且两种情况均是ACC的含水形式。也就是说,ACC是生物矿化作用[17,31]所涉及到的一种特殊的钙碳酸盐的矿物形式[27],是一种较为典型的生物矿物的过渡型前体相的矿物形式[31,32]。一个基本事实是,生物矿物一直到新元古代最晚期才开始出现[33];穿越前寒武纪到寒武纪,沉积面和沉积物内发生的重要变化是生物扰动作用和与生物矿化作用相关的“生物骨骼化作用”,这个变化还被誉为“生物历史性的底栖农艺革命”[34,35]。
如果形成鲕粒皮层的ACC微薄层,可理解为一种含水的而且含有有机大分子矿物相的话,Duguid等[1]所描述的现代鲕粒的形成似乎与微生物活动存在一种复杂的联系;作为生物矿化(或生物骨骼化)作用的过渡前体矿物相的ACC可以形成在生物体外[27],这种形成鲕粒皮层的ACC可能代表了“非酶控制的碳酸钙沉淀作用”,即产生有机矿化作用的有机矿物[17],这种不但含水而且含有有机大分子的非晶质钙碳酸盐(ACC)也能由微生物活动产生,尤其是在蓝细菌分泌的细胞外聚合物质之中[36~39];参与有机矿化作用的有机高分子是聚阴离子的聚合物,这些聚合物分开那些由诸如氨基酸和糖的单体组成的分子链。由于没有考虑到Reitner和Arp等的研究成果[36~39],还不能将Duguid等[1]所描述的现代鲕粒形成的化学过程理解为一个纯粹的化学作用模式,因为形成鲕粒皮层的ACC可能是蓝细菌分泌的EPS中的有机矿物;实际上,Duguid等[1]所描述的现代鲕粒中,大量的蓝细菌产生的微生物钻孔也间接地说明了形成鲕粒皮层的ACC可能与蓝细菌活动有关。
4 华南下三叠统的鲕粒
湖北利川剖面三叠系大冶组顶部的鲕粒灰岩[12],形成在早三叠世印度期上扬子区鲕粒滩自西向东进积作用过程的晚期[40,41],即一个缓坡型台地的生长发育过程之中,而该缓坡型台地正好形成在中国南方二叠纪-三叠纪之交的大规模台地淹没事件之后。形态多样的鲕粒(如图1所示),特别是显生宙较为罕见的巨鲕的发育,反映了二叠纪末期生物大灭绝事件之后特殊的荒凉海底环境的沉积作用特征,从而代表了一种特殊的“时错相”[42~45]。
下三叠统的鲕粒,可以见到大于2 mm、甚至可以达到7~8 mm直径的“巨鲕”(图1的A,B所示),这是自从在新元古界发现的“巨鲕”[46,47]之后,在显生宙所发现的较为罕见“巨鲕”[12]。最为特别的是,这些直径常常大于2 mm的鲕粒,表现出以下具有“生物活力”的现象,而支持了Brehm等[10,11]所提出的“球状微生物群落”形成生物微球、生物微球形成鲕粒的微生物作用学说。这些具有明显“生物活力”的沉积学现象表现在以下两个方面:
第一、不管是何种形态的鲕粒,包括圆形同心鲕粒(图1A)、椭圆形同心鲕粒(图1B)、复合鲕粒(图1C)和偏心鲕粒(图1D),鲕粒的皮层均表现出明显的“亮层”和“暗层”,显示出类似于叠层石的特点,只不过是球形叠层石而已,区别于那些形成于垂向排列的纹层状叠层石。考虑到叠层石是有名的形成于垂向排列的微生物膜或微生物席的纹层状岩石[13,15],而且叠层石作为“微生物席的建造物”[7,11,13~17,36~39,47~49]已经成为共识,这个现象似乎更多地支持了Brehm等[10,11]所提出的“球状微生物群落”形成鲕粒的假说。
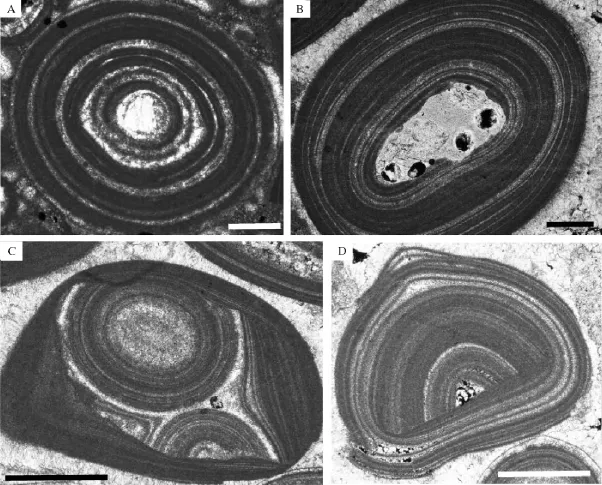
图1 湖北利川三叠系大冶组的鲕粒A、圆形同心鲕粒;B、椭圆形同心鲕粒;C、复合同心鲕粒;D、偏心鲕粒.比例尺为1 mmFig.1 Oolites from the Daye Formation of the lower Triassic at the Lichuan section in Hubei Province A,Circular-shaped concentric oolite;B,elliptical-shaped concentric oolite;C,composite concentric oolite;D,eccentric oolite.Scale bars,1 mm
第二、如图1C和1D所示,那些复合鲕粒和偏心鲕粒,显示出明显的“再生”特征和“自我修复”的能力,从而显示出明显的“生物活力”。就像叠层石本身不能作为生物体的代表一样,这种再生特征和自我修复的能力所表现出的“生物活力”,难以用纯粹的物理作用和化学作用过程来进行解释,尽管在图1所示的化石鲕粒之中已经找不到直接的微生物作用证据,还是间接地说明了鲕粒的形成与微生物活动存在关联,从而也支持了Brehm等[10,11]所提出的“球状微生物群落”形成鲕粒的假说。
也可能正是形成在动荡的高能环境之中,图1所示的浮游鲕粒围绕一个核心颗粒的、而且连续不断的同心层聚集作用来抗拒强烈的水流破坏和改造,明显的亮层和暗层似乎意味着这些纹层就像叠层石一样是微生物席或微生物膜的残余物,即特殊的球状微生物群落的产物[10,11],如果水流的动荡程度降低一些有可能这些纹层会变成扰动的纹层,从而形成球状叠层石——核形石。这种在强烈水流作用下的微生物作用过程的推测,还需要进一步研究才能得到合理的诠释。
5 华北寒武系崮山组的鲕粒
在新的寒武系年代地层系统[50~53]中,华北地台北缘寒武系第三统中上部为一个较为典型的鲕粒滩相灰岩组成的地层序列[54,55],大致相当于徐庄组、张夏组和崮山组的顶部,分别组成三套较为典型的鲕粒滩相灰岩地层[54,56~60]。其中,最有代表性的剖面是北京西郊的下苇甸剖面,该剖面的崮山组构成一个三级层序,三级层序的高水位体系域正是由鲕粒滩相灰岩地层所组成[54]。如图2所示,在崮山组上部的高水位体系域鲕粒滩相灰岩中,颗粒滩相灰岩本身由浮游鲕粒所组成,在其中呈串珠状发育的叠层石生物丘中还发育着一些特殊的底栖鲕粒,从而成为窥视和了解底栖鲕粒和浮游鲕粒[61]的典型地点。

图2 北京西郊下苇甸剖面寒武系崮山组上部的块状鲕粒滩相灰岩及其中的叠层石生物丘A.呈串珠状分布在块状鲕粒滩灰岩中的叠层石生物丘(箭头所指);B.叠层石生物丘发育的柱状叠层石; C.生物丘的顶界(箭头所指);D、生物丘的左侧边界(箭头所指)Fig.2 Images showing the stromatolitic biohermswithin themassive bank-facies oolitic grainstones in the upper part of the Cambrian Gushan Formation at the Xiaweidian section in the western suburb of Beijing A.stromatolitic bioherms(the arrowed)that are distributed in the string of beads within themassive oolitic grainstones of bank facies; B.column stromatoilites developed within bioherm;C,the top boundary(the arrowed);B:the left boundary of the stromatolitic bioherm (the arrowed);C.the right boundary(the arrowed)of the stromatolitic bioherm
在中奥陶世生物大辐射事件[62]之前的寒武纪至早奥陶世,骨骼还没有成为全球持续性的碳酸盐沉积物的重要来源,所以该时期的海洋被称为“贫乏骨骼的海洋”[63];也由于该时期在全球范围的大部分地层之中总是发育特殊的以竹叶状砾屑灰岩为代表的风暴沉积,所以该时期还被誉为“风暴海”[64~67]。在处于显生宙早期的这个承前启后的特殊地质时期,随着后生动物在寒武纪期间的辐射而微生物碳酸盐岩也在增加,形成了显生宙早期的第一幕蓝细菌钙化作用事件[49],并进一步称为微生物碳酸盐岩的“寒武纪-早奥陶世复苏期”[68,69]。在寒武纪早期的古杯海绵骨骼生物礁生态系崩溃之后的寒武纪第三世至芙蓉世,进入了一个后生动物骨骼生物礁贫乏的特殊时代而发育微生物礁的时代[70~74];从更宽泛的涵义上讲[75],图2所示的圆丘状叠层石生物丘,普遍发育的叠层石表明这种块状碳酸盐构造也可以归入到“微生物礁”的范畴。
5.1 浮游鲕粒
与图1所示的下三叠统的颗粒滩相鲕粒一样,寒武系崮山组的颗粒滩相鲕粒则组成较为典型的浮游鲕粒(pelagic ooid)[2,3,61,76]。这些可以归为颗粒滩相灰岩的厚层块状鲕粒灰岩,其鲕粒主要为0.5~1.0 mm大小的圆形和次圆形泥晶-放射鲕粒,颗粒之间的填充物多为泥晶,局部为亮晶胶结物(图3A和B所示)。尽管以放射状-泥晶鲕粒为特征(图3A至C),这些鲕粒较为均匀的颗粒大小、以及多呈厚层或块状产出的特点(图2A),属于浅海高能颗粒滩相沉积的浮游鲕粒[2,3,61,76]。
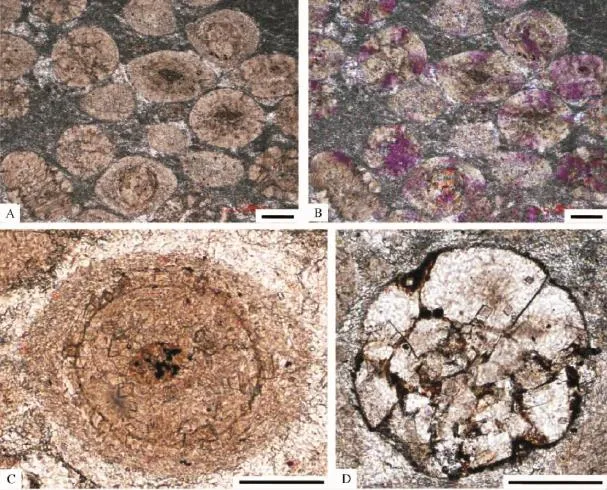
图3 北京西郊下苇甸剖面寒武系崮山组上部颗粒滩相灰岩中的浮游鲕粒A和B.多为放射-同心鲕粒组成的颗粒滩相灰岩,A为单偏光照片,B为正交光照片;C.典型的放射-同心鲕粒,单偏光照片; D.由黑色富有机质的泥晶粘聚而成的假鲕粒,单偏光照片.比例尺为100μmFig.3 Images showing the pelagic ooids in the upper part of the Cambrian Gushan Formation at the Xiaweidian section in the western suburb of Beijing A and B,bank-facies oolitic grainstonesmainly composed of concentric-radiated oolites,A is the image under themonopolarized light,and B is the image under the crossed-polarized light;C delegates one enlarged concentric-rediated ooid under themonopolarized light;D is amagnified pseudo-oolitemade up of echinoderm bioclasts bound by dark micrites enriched with organic substances under themonopolarized light.Scale bars are 100 microns
这些构成颗粒滩的高能浮游鲕粒,与上文的图1所示的下三叠统高能浮游鲕粒不同的是多为放射纤维状方解石所组成,鲕粒的不太明显的圈层多为黑色泥晶构成(图3A至D);也就是说,类似于Duguid等[1]所描述的现代巴哈马的鲕粒,尽管经过了成岩作用的改造,这些浮游鲕粒多为非晶质钙碳酸盐(ACC)所组成,那些不太明显的圈层似乎是富有机质的细菌泥晶[76]所构成,如果不考虑这些表现为放射纤维状方解石的ACC来源的话,崮山组中的这些浮游鲕粒可以运用Duguid等[1]所提出的化学作用机制来加以解释。但是,如图3D所示,除了泥晶-放射状鲕粒以外,崮山组上部的颗粒滩相鲕粒灰岩中还发育圆形假鲕粒,这种假鲕粒由富含有机质的黑色泥晶(细菌成因的碳酸盐泥晶[77~81])粘聚棘皮生物碎屑所形成,不但增加了这些浮游鲕粒的多样性,而且也说明了这些悬浮鲕粒直接或间接地与微生物活动存在一些联系,因为这些黑色富有机质的隐晶质碳酸盐泥晶,可能是石内蓝细菌产生的微生物钻孔过程所诱发的泥晶化作用的产物[77~81]而进一步归为细菌泥晶。这些浮游鲕粒,从圆形到椭圆形乃至不规则的形态,有的具核心,有的不见核心(图3A至C);如果考虑到那些表现为放射纤维状方解石的ACC可能是与蓝细菌或其他微生物活动相关的分泌物的话[36~39],再考虑到微生物钻孔过程相关的泥晶化作用[77~81]所形成的富有机质的黑色泥晶形成的鲕粒圈层,就像细菌泥晶粘结棘皮类生物碎屑所形成的类似于滚雪球式的假鲕粒(图3D所示),图3A至C所示的浮游鲕粒的形成直接地或间接地与微生物活动存在成因关联,更多地支持了Brehm等[10,11]的鲕粒形成的微生物作用假说。
5.2 底栖鲕粒
发育在崮山组上部的柱状叠层石生物丘,在显微镜下,这些柱状叠层石(图2B所示)总体上表现为致密泥晶结构,属于较为典型的“泥晶相叠层石”[47,82~84](图4所示);这些柱状叠层石组成的圆丘状构造,产在高能鲕粒滩相灰岩之中,又与前人所描述的寒武纪深水叠层石生物礁[70,71,85,86]存在一些差异。在崮山组叠层石生物丘内部,发育放射-纤维状方解石(或文石?)为主所构成的各种形态和微组构的底栖鲕粒[2,3,61,76](图4),这些多为漂浮状产出在致密隐晶质泥晶组成的叠层石内部的鲕粒,特殊的产出背景明显不同于与Reitner等[36]所描述的美国犹他州大盐湖中的放射鲕粒。如果将叠层石理解为微生物席的建造物[7,11,13~17,36~39,47~49]的话,崮山组叠层石生物丘中的鲕粒,就成为Gerdes等[61]所描述的微生物席之中生物成因的碳酸盐颗粒多样性的一个具有代表性的化石实例。之所以称之为底栖鲕粒,是因为他们缠在叠层石内部,不是浮游在在水体之中而区别于浮游鲕粒。
关于微生物活动所控制的碳酸盐沉淀作用[87~92],涉及到细菌降解和有机质的转变作用过程之中复杂的生物化学反应,Krumbein[93,94]曾经总结了一系列钙化作用的生物控制作用机理,包括黑暗环境的二氧化碳固定作用、厌氧呼吸作用和发酵作用、二氧化碳的吸收和转变作用、氨的生产作用、碱度和pH的变化,以及无机磷形成过程的细菌调节作用[95,96]。一些学者曾经采用碳酸盐沉积学的术语来描述沉淀在微生物席中的单个颗粒形式,如核形石、鲕粒和葡萄石[97,98],以及被假定为结晶作用的非排泄过程所形成的球粒[99]等。除了钙碳酸盐矿物以外,其他的矿物形式也能形成颗粒,如磷质包覆颗粒的形成[100,101],以及微生物席内铁质颗粒的形成[102]。前人的这些研究为分析崮山组叠层石生物丘内的底栖鲕粒的形成提供了极为重要的线索和理论基础。
崮山组叠层石生物丘中的底栖鲕粒[2,3,61,76],颗粒直径多为20~50μm,一般不超过50μm,颗粒直径明显小于围岩中的鲕粒滩相灰岩中的浮游鲕粒(图3所示)。这些鲕粒多为圆形,少数为次圆性或不规则形状,鲕粒圈层数目较少,有核心或没有核心,鲕粒内部残留着较多的有机质残余物和少量泥晶;根据主要形态、核心和鲕粒圈层的是否发育等,大致可以划分为有核放射-同心鲕粒、无核放射-同心鲕粒、有核放射鲕粒、无核放射鲕粒、不规则形态的鲕粒等等(图4),从而显示出在叠层石所表征的微生物席内底栖鲕粒的形态多样性。
如图4所示,这些形态各异、有核心和没有核心的放射状方解石(或文石)组成的底栖鲕粒,表现了以下几个特点:1)鲕粒的外部圈层多表现为泥晶套(图4A,B和D,F所示),这种泥晶套厚度为数微米至数十微米不等,有时还显示出类似叠层石纹层的球形纹理(图4D所示);2)鲕粒的同心圈层平滑但不连续,厚度数微米不等,与鲕粒外部圈层的泥晶套一样主要为隐晶质泥晶所组成,可能代表了微生物膜的残余物[61];3)沿着鲕粒的同心圈层以及鲕粒外部圈层的泥晶套,总是发育垂直于鲕粒核心或圈层的小黑点(图4C,F所示),它们类似于前人描述和解释的、石内蓝细菌产生的微生物钻孔的残余物[1,77~81],这些可能的微生物钻孔残余物有些还表现为长数微米、直径为1~2μm的小管,多垂直于鲕粒核心展布;4)虽然还难以确切知道构成这些底栖鲕粒的放射-纤维状方解石(或文石?)的物质来源和形成机制,但是,就像Gerdes等[61]指出的那样,这些纤维状方解石(或文石?)的结壳作用与形成鲕粒圈层的微生物膜的交互作用是形成这种鲕粒的主要过程。如果把构成这些底栖鲕粒的放射-纤维状方解石(或文石?)解释为非晶质钙碳酸盐(ACC),而且是微生物席内的各种微生物活动如硫酸盐还原反应、异养细菌的呼吸作用等微生物新陈代谢活动所形成的产物的话,更进一步讲,这些微亮晶有可能是叠层石代表的微生物席内的含水而且富有机质(主要为细胞外聚合物质)的介质之中直接的沉淀作用产物[48,49,103,104],表明了这些底栖鲕粒的形成和生长与复杂的微生物新陈代谢活动存在着较为紧密的成因联系。因此,崮山组叠层石生物丘中的底栖鲕粒,特殊的物质构成(放射-纤维状方解石(或文石)组成的ACC)、以及特殊的微组构(受到微生物钻孔改造、以及由其所造成的泥晶化作用产生的球形微生物膜残余物组成的平滑但有不连续的鲕粒圈层),明显地支持了这些底栖鲕粒的微生物成因[10,11,61]。
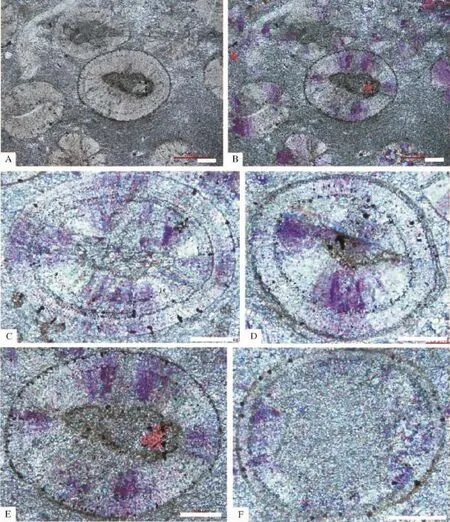
图4 北京西郊下苇甸剖面崮山组上部叠层石生物丘中的底栖鲕粒A和B.发育底栖鲕粒的泥晶相叠层石的显微组构,A为单偏光,B为正交光;C.具核心的椭圆形同心放射鲕粒;D.具核心的圆形同心放射鲕粒; E.具核心的放射鲕粒;F.无核心的放射薄皮鲕粒.C至F为正交光照片,比例尺均为100μmFig.4 Images showing the benthic ooids within stromatolitic bioberms in the upper part of the Cambrian Gushan Formation at the Xiaweidian section in the western suburb of Beijing A and B refer to the generalmicrofabric of the stromatolites of themicritic facies,A the image under themonopolarized light and B is the image under the crossed-polarized light;C,elliptical radiated-concentric oolite with nucleus;D,circular radiated-concentric oolite with nucleus;E,elliptical radiated oolite with nucleus;F,circular radiated oolite without nucleus.C to F are images under the crossed-polarized light,scale bars are 100 microns
6 结语和讨论
作为最为迷人的碳酸盐岩的一种包覆颗粒的类型,鲕粒的成因一直是困扰沉积学家的一个难题之一。近年来,形成了两种对立的观点,第一种是在Gerdes等[61]研究基础上建立起来的鲕粒形成的微生物成因学说,第二种是Duguid等[1]的新的化学作用学说。
来自于华南下三叠统的鲕粒,不但发育着显生宙罕见的“巨鲕”[12],而且这些巨鲕所具有的类似于叠层石的组构、以及再生现象和自我修复现象表现出的“生物活力”,间接地支持了鲕粒形成的微生物作用机制;华北寒武系崮山组的浮游鲕粒,以及叠层石生物丘中的底栖鲕粒,主要由表现为放射-纤维状方解石(或文石?)的ACC所组成,底栖鲕粒中也见到明显的微生物钻孔残余物,说明鲕粒形成与微生物活动的紧密联系。当然,还不能将鲕粒简单地归为“微生物碳酸盐岩”[13,49],因为不同组构鲕粒发育环境的多样性又说明其形成与形成环境的化学条件和物理条件的紧密而且复杂的联系;只能说,鲕粒形成的微生物过程可能是一个形成鲕粒的主要作用机制。但是,就像Dupraz等[48]强调的那样,微生物新陈代谢作用和其他的物理化学属性之间的协同作用对于微生物生态系在地球历史中的成功起到了关键作用,这就需要在不同种群之间或者其内存在一个化学交换,所有这些过程都需要具有一个详细的了解,而现在我们只是触及到“碳酸盐岩冰山”的一角;更进一步讲,最大的挑战是所有这些过程和产物在进行岩石记录的解释时,必须穿越成岩作用“过滤器”,许多原生的沉积信息已经损失在漫长的成岩作用过程之中。随着研究的更加深入,鲕粒形成的复杂过程所代表的“有机矿化作用”机制将会取得更加清晰的阐释,也希望本文的介绍和评述能够起到抛砖引玉的作用,而为今后更加深入的研究提供更多有益的思考途径。
References)
1 Duguid SM A,Kyser T K,James N P,etal.Microbes and ooids[J].ournal of Sedimentary Research,2010,80:236-251
2 Simone L.Ooids:a review[J].arth-Science Reviews,1980,16: 319-355
3 Davies,P J,Bubela B,Ferguson,J.The formation of ooids[J].edimentology,1978,25:70-729
4 Gaffey S J.Formation and infilling of pits inmarine ooid surfaces[J].ournal of Sedimentary Petrology,1983,53:193-208
5 Folk R L,Lynch F L.Organicmatter,putative nannobacteria and the formation of ooids and hardgrounds[J].edimentology,2001,48: 215-229
6 Fabricius F H.Origin of Marine Ooids and Grupestones[M]∥Stuttgart:E.Schweizerbart'sche Verlagsbuchhandlung(Nagele u.Obermiller),1977:1-113
7 Reid R P,Visscher P T,Decho A W,et al.The role ofmicrobes in accretion,lamination and early lithification ofmodernmarine stromatolites[J].ature,2000,406:989-992
8 Newell N D,Purdy E G,Imbrie J.Bahamanian o litic sands[J].ournal of Geology,1960,68:481-497
9 Tucker M E,Wright V P.Carbonate Sedimentology[M].xford: Blackwell Sciences,1990
10 Brehm U,Krumbein W E,Palinska K A.Microbial spheres:a novel cyanobacterial-diatom symbiosis[J].aturwiss,2003,90:136-140
11 Brehm U,Krumbein W E,Palinska K A.Biomicrospheres generate ooids in laboratory[J].eomicrobiology Journal,2006,23:545-550
12 梅冥相。显生宙罕见的巨鲕及其鲕粒形态多样性的意义:以湖北利川下三叠统大冶组为例[J]。现代地质,2008,22(5):683-698 [MeiMingxiang.Implication for the unusual giantoolites of the Phanerozoic and theirmorphological diversity:a case study from the Triassic Daye Formation at the Lichuan section in Hubei Province,South China[J].eoscience,2008,22(5):683-698]
13 梅冥相。微生物碳酸盐岩分类体系的修订:对灰岩成因-结构分类体系的补充[J]。地学前缘,2007,14(5):597-614[Mei Mingxiang.Revised classification ofmicrobial carbonates:Replenishment to the classification of limestones[J].arth Science Frontiers, 2007,14(5):597-614]
14 Krumbein W E.Stromatolities-the challenge of a term in space and time[J].recambrian Research,1983,20:493-531
15 Altermunn W.Precambrian stromatolites:Problems in definition, classification,morphology and stratigraphy[C]∥Eriksson PG,Altermann W.Nelson D R,etal.The Precambrian Earth:Tempos and E-vents.Amsterdam:Elsevier,2004:564-574
16 Pinckney JL,Reid R P.Productivity and community composition of stromatolitic microbialmats in the Exuma Cays,Bahamas[J].acies,1997,36:204-207
17 Perry R S,Mcloughlin N,Lynne B Y,et al.Defining biominerals and organominerals:Direct and indirect indicators of life[J].edimentary Geology,2007,201:157-179
18 Zavarzin G A.Microbial geochemical calcium cycle[J].icrobiology (Mosscow,English Translation),2002,71:1-17
19 Hammes F,Boon N,De Villiers J,et al.Strainspecific ureolyticmicrobial calcium carbonate precipitation[J].pplide and Environmental Microbiology,2003,69:4901-4909
20 Contos A K,James JM,Heywood B,etal.Morphoanalysis of bacterially precipitated subaqueous calcium carbonate from Weebubbie Cave,Australia[J].eomicrobiology Journal,2001,18:331-343
21 Davaud E,Girardclos S.Recent freshwater ooids and oncoids from Western Lake Geneva(Switzerland):indications of a common organi-callymediated origin[J].ournal of Sedimentary Research,2001, 71:423-429
22 Mitterer R M.Amino acid composition of organicmatrix in calcareous oolites[J].cience,1968,162:1498-1499
23 Mitterer R M.Biogeochemistry of aragonite mud and oolites[J].eochimica et Cosmochimica Acta,1972,36:1407-1422
24 Folk R L.SEM imaging of bacteria and nannobacteria in carbonate sediments and rocks[J].ournal of Sedimentary Petrology,1993, 63:990-999
25 Folk R L,Lynch F L.Organicmatter,putative nannobacteria and the formation of ooids and hardgrounds[J].edimentology,200,48: 215 229
26 Romanek C S,Grossman E L,Andmorse JW.Carbon isotopic fractionation in synthetic aragonite and calcite:Effects of temperature and precipitation rate[J].eochimica et Cosmochimica Acta,1992,56: 419-430
27 Addadi L,Rax S,Weiner S.Taking advantage of disorder:Amorphous calcium carbonate and its roles in biomineralization[J].dvanced Materials,2003,15:959 970
28 Schmidt M,Xeflide S,Botz R,et al.Oxygen isotope fractionation during synthesis of Ca Mg-carbonate and implications for sedimentary dolomite formation[J].eochimica et Cosmochimica Acta,2005, 69:4665 4674
29 Morse JW,Gorman B P,Lee J,et al.Ingerson lecture:Nanoscale insights into the mechanisms responsible for the strange kinetic and solubility behaviour of aragonite in seawater[J].eological Society of America,Abstractswith Programs,2008,40:457-459
30 Loreau JP,Purser B H.Distribution and Ultrastructure of Holocene Ooids in the Persian Gulf[C]∥Purser B H.The Persian Gulf:Holocene Carbonate Sedimentation and Diagenesis in a Shallow Epicontinental Sea.New York:Springer-Verlag,1973:279-328
31 Mann S.Biomineralization-Principles and Concepts in Bioinorganic Materials Chemistry[M].xford:Oxford University Press,2001
32 Skinner H C W,Jahren A H.Biomineralization:Treatise on geochemistry[J].msterdam:Elsevier,2003:117 184
33 Knoll A H.Biomineralization and evolutionary history[C]∥Dove P M,Yoreo J J,Weiner S.Biomineralization:Reviews in Minerolgy and Geochemistry,2003,54:329-356
34 Seilacher A,Pflügel E.From biomats to benthic agriculture:A biohistoric revolution[C]∥Krumbein W E,Paterson D M,Stal L J.Biostabilization of Sediments.Oldenberg:Bibliotheks und Informationssystem der Carl von Ossietzky Universit t Oldenberg,1994:97-105
35 Seilacher A.Biomat-related lifestyles in the Precambrian[J].alaios,1999,14:86-93
36 Reitner J,Arp G,Thiel V,et al.Organic matter in Great Salt Lake Ooids(Utah,USA)Firstapproach to a formation via organicmatrices [J].acies,1997,36:210-219
37 Reitner J,Thiel V,Zankl H,et al.Organic and biochemical pattern in cryptic microbialites[C]∥Riding R,Awramik S M.Microbial Sediments.Berlin:Springer,2000:149-160
38 Arp G,Reimer A,Reitner J.Photosynthesis-induced biofilm calcification and calcium concentrations in Phanerozoic Ocean[J].cience, 2001,292:1701-1704
39 Arp G,Reimer A,Reitner J.Microbialite formation in seawater of increased alkalinity,Satonda Crater Lake,Indonesia[J].ournal of Sedimentary Research,2003,73:105-127
40 陈公信,金经炜。湖北省岩石地层[M]。武汉:中国地质大学出版社,1997[Chen Gongxin,Jin Jingwei.Lithostratigraphy in Hubei Province[M].uhan:China University of Geosciences,1997]
41 吴应林,朱洪发,朱忠发,等。中国南方三叠纪岩相古地理与成矿作用[M]。北京:地质出版社,1994[Wu Yinglin,Zhu Zhongfa, Zhu Zhongfa,etal.Triassic Lithofacies Palaeogeography and Mineralization in South China[M].eijing:Geological Publishing House, 1994]
42 赵小明,童金南,姚华舟,等。华南早三叠世错时相沉积及其对复苏期生态系的启示[J]。中国科学D辑:地球科学,2008,38 (12):1564-1574[Zhao Xiaoming,Tong Jinnan,Yao Huazhou,et al.Anachronistic facies in the Lower Triassic of South China and their implications to the ecosystems during the recovery time[J].cience in China Series D:Earth Sciences,2008,51(11):1646-1657]
43 李飞,王夏,薛武强,等。一种新的错时相沉积物:巨鲕及其环境意义[J]。沉积学报,2010,28(3):585-595[Li Fei,Wang Xia,XueWuqing,et al.Origin and environmental significance of giant ooids in the Early Triassic:a new kind of anachronistic facies[J].cta Sedimentologica Sinica,2010,28(3):585-595]
44 赵小明,牛志军,童金南,等。早三叠世生物复苏期的特殊沉积: “错时相”沉积[J]。沉积学报,2010,28(2):314-323[Zhao Xiaoming,Niu Zhijun,Tong Jinnan,etal.The distinctive sediments in the Early Triassic recovery time:“Anachronistic Facies”[J].cta Sedimentologica Sinica,2010,28(2):314-323]
45 Pruss SB,Corsetti F A,Bottjer D J.The unusual sedimentary rock record of the Early Triassic:a case study from the southwestern United States[J].alaeogeography,Palaeoclimatology,Palaeoecology, 2005,222:33-52
46 Summer D A,Grotzinger JP.Numericalmodeling of ooid size and the problem of Neoproterozoic giantooids[J].ournal of Sedimentary Petrology,1993,63:974-982
47 Grotzinger JP,James N P.Precambrian carbonates:evolution of understanding[C]∥Grotzinger,JP,James N P.Carbonate Sedimentation and Diagenesis in the Evolving Precambrian World.SEPM Special Publication 67,2000:3-22
48 Dupraz C,Reid R P,Braissant O,et al.Processes of carbonate precipitation in modern microbial mats[J].arth-Science Reviews, 2009,96:141-162
49 Riding R.Microbial Carbonates:The geological record of calcified bacterial-algal mats and biofilms[J].edimentology,2000,47 (Suppl。1):179-214
50 彭善池,Bobcock L E。全球寒武系年代地层再划分的建议[J]。地层学杂志,2005,29(1):92-93,96[Peng Shanchi,Bobcock L E.Newly proposed global chronostratigraphy subdivision on Cambrian System[J].ournal of Stratigraphy,2005,29(1):92-93,96]
51 彭善池。全球寒武系四统划分框架正式确立[J]。地层学杂志, 2006,30(2):147-148[Peng Shanchi.A new global framework with four series for Cambrian system[J].ournal of Stratigraphy,2006,30 (2):147-148]
52 彭善池。华南寒武系年代地层系统的修订及相关问题[J]。地层学杂志,2008,32(3):239-245[Peng Shanchi.Revision on Cam-brian chronostratigraphy of South China and related remarks[J].ournal of Stratigrapphy,2008,32(3):239-245]
53 章森桂,张允白,严慧君,2009。“国际地层表”(2008)简介[J]。地层学杂志,2009,33(1):1-10[Zhang Senggui,Zhang Yunbai,Yan Huijun.A brief introduction to the'International Stratigraphic Chart'(2008)[J].ournal of Stratigraphy,2009,33(1): 1-10]
54 梅冥相,马永生,梅仕龙,等。华北寒武系层序地层格架及碳酸盐台地演化[J]。现代地质,1997,11(3):275-282[Mei Mingxiang,Ma Yongsheng,Mei Shilong,et al.Sequence-stratigraphic framework and carbonate-platform evolution for the Carmbrian of the North-China Platform[J].eoscience,1997,11(3):275-282]
55 Meng Xianghua,Ge Ming,Tucker M E.Sequence stratigraphy,sealevel changes and depositional systems in the Cambro-Ordovician of the North China carbonate platform[J].edimentary Geology,1997, 114:189-222
56 柳永清,孟祥化,葛铭。华北地台中寒武世鲕滩碳酸盐旋回沉积、古海平面变动控制及旋回年代学研究[J]。地质科学,1999, 34(4):442-450[Liu Yongqing,Meng Xianghua,Ge Ming.The sealevel change forcing cycles of oolitic carbonate and cyclochrological applications[J].cientia Geologica Sinica,1999,34(4):442-450]
57 张旭,张宁,杨振鸿,等。北京西山下苇甸中寒武统碳酸盐岩微相及沉积相研究[J]。地质科技情报,2009,28(6):27-30[Zhang Xu,Zhang Ning,Yang Zhenhong,et al.Carbonate Microfacies and sedimentary faciesofmiddle Cambrian Formation at Xiaweidian Profile in Western Hills,Beijing,China[J].eological Science and Technology Information,2009,28(6):27-30]
58 梅冥相,梅仕龙。华北中寒武世张夏组复合海平面变化旋回层序[J]。沉积学报,1997,15(4):5-10[MeiMingxiang,Mei Shilong.Cyclic-sequences of composite sea-level change developed in Zhangxia Formation of Middle-Cambrian in North-China[J].cta Sedimentologica Sinica,1997,15(4):5-10]
59 章雨旭。华北板块寒武系的穿时性[J]。沉积与特提斯地质, 2001,21(1):78-87[Zhang Yuxu.Diachronism of Cambrian strata on the North China Platform[J].edimentary Geology and Tethyan Geology,2001,21(1):78-87]
60 孟祥化,乔秀夫,葛铭。华北古浅海风暴沉积及丁家滩相序模式[J]。沉积学报,1986,5(2):1-18[Meng Xianghua,Qiao Xiufu and Ge Ming.Storm deposits of the paleo-epicontinental sea and the Dingjiatan model in the Cambrian of the North China Platform[J].cta Sedimentologica Sinica,1986,5(2):1-18]
61 GerdesG,Dunajtschik-Piewak K,Riege H,etal.Structural diversity of biogenic carbonate particles in microbialmats[J].edimentology, 1994,41:1273-1294
62 Sepkoski J J.Biodiversity:Past,present,and future[J].ournal of Paleontology,1997,71:533-539
63 Pruss SB,Finnegan S,FischerW W,et al.Carbonates in skeletonpoor seas:new insights from Cambrian and Ordovician strata of Laurentia[J].alaios,2010,25:73 84
64 Sepkoski J J.Flat-pebble conglomerates,storm deposits,and the Cambrian bottom fauna[C]∥Einsele G,Seilacher A.Cyclic Event and Stratification.Berlin:Springer-Verlag,1982:371 388
65 Pratt BR.Storms versus tsunamis:Dynamic interplay of sedimentary, diagenetic,and tectonic processes in the Cambrian of Montana[J].eology,2002,30:423-426
66 Pratt BR,Bordonar O L.Tsunamis in a stormy sea:middle Cambrian inner-shelf limestones of western Argentina.Journal of Sedimentary Research,2007,77:256-262
67 Myrow PM,Tice L,Archuleta B,et al.Flat-pebble conglomerate: itsmultiple origins and relationship tometre-scale depositional cycles [J].edimentology,2004,51:973-996
68 Riding R,Liang L.Geobiology ofmicrobial carbonates:metazoan and seawater saturation state influences on secular trends during the Phanerozoic[J].alaeogeography,Palaeoclimatology,Palaeoecology, 2005,219:101-115
69 Riding R.Microbial carbonate abundance compared with fluctuations in metazoan diversity over geological time[J].edimentary Geology, 2006,185:229-238
70 Wood R.The changing biology of reef-building[J].alaios,1995, 10:517-529
71 Wood R.Reef Evolution[M].xford:Oxford University Press,1999
72 Webb G E.Was Phanerozoic reef history controlled by the distribution of non-enzymatically secreted reef carbonates(microbial carbonates and biologically induced cement)?[J].edimentology,1996,43: 947-971
73 KiesslingW.Paleoclimatic significance of Phanerozoic reefs[J].eology,1999,29:751-754
74 KiesslingW,Flügel.E,Golonka J.Paleoreefmaps:evaluation of a comprehensive database on Phanerozoic reefs[J].APG Bulletin, 1999,83:1552-1587
75 Riding R.Structure and composition of organic reefs and carbonate mud mounds:concepts and categories[J].arth-Science Reviews, 2002,58:163-231
76 Flügel.E.Microfacies of Carbonate rocks:Analysis,Interpretation and Application[M].erlin:Springer-Verlag,2004
77 Perry C T.Grain susceptibility to the effects ofmicroboring:implications for the preservation of skeletal carbonates[J].edimentology, 1998,45,39-51
78 Perry C T,Macdonald IA.Impacts of light penetration on the bathymetry of reefmicroboring communities:implications for the development of microendolithic trace assemblages[J].alaeogeography Palaeoclimatology Palaeoecology,2002,186:101-113
79 Golubic S,Seong-Joo L,Browne K M.Cyanobacteria:architects of sedimentary structures[C]∥Riding R E,Awramik SM.Microbial Sediments.Berlin:Springer-Verlag,2000,57-67
80 Garcia-Pichel F.Plausiblemechanisms for the boring on carbonatesby microbial phototrophs[J].edimentary Geology,2006,185:205-213
81 Chac n E,Berrendero E,Pichel FG.Biogeological signatures ofmicroboring cyanobacterial communities in marine carbonates from Cabo Rojo,Puerto Rico[J].edimentary Geology,2006,185:215-228
82 Kah L C,Knoll A H.Microbenthic distribution of Proterozoic tidal flats:Environmental and taphonomic considerations[J].eology, 1996,24:79-82
83 梅冥相,高金汉,孟庆芬,等。天津蓟县中元古界雾迷山组微指状叠层石及其对1250Ma±叠层石衰减事件的响应[J]。古地理学报,2008,10(5):495-509[Mei Mingxiang,Gao Jinhan,Meng Qingfen,et al.Microdigital stromatolites and their response to the stromatolite decline at ca。1250 Ma for the Mesoproterozoic Wumishan Formation at Jixian section in Tianjin[J].ournal of Palaeogeography,2008,10(5):495-509]
84 MeiMingxiang,Gao Jinhan,Meng Qingfen,et al.Sedimentary features and their implications ofmicrodigital stromatolites from theMesoproterozoic Wumishan Formation at the Jixian section in North China [J].cta Geologica Sinica(English Edition),2010,82(3):483-496
85 Wood R.Are reefs and mud mounds really so different?[J].edimentary Geology,2001,145:161-171
86 Pratt B R.Microbial contribution to reefal mud-mounds in ancient deep-water settings:evidence from the Cambrian[C]∥Riding R, Aramik SM.Microbial Sediments.Berlin:Springer-Verlag,2000, 282-293
87 KrumbeinW E.On the precipitation ofaragonite on the surface ofmarine bacteria[J].aturwissenschaften,1974,61:167-178
89 Buczynski C,Chafetz H S.Habitofbacterially induced precipitates of calcium carbonate and the influence ofmedium viscosity onmineralogy [J].ournal of sedimentary Petrology,1991,61:226-233
90 Chafetz H S,Rush P F,Utech N M.Microenvironmental controls on mineralogy and habit of CaCO3,precipitates:an example from an active travertine system[J].edimentology,1991,38:107-126
91 Chafetz H S,Bucaynski C.Bacterially induced lithification ofmicrobialmats[J].alaios,1992,7:277-293
92 Riege H,Gerdes G,Krumbein W E.Contribution of heterotrophic bacteria to the formation of CaCO3aggregates in hypersalinemicrobial mats[J].ieler Meersforschungen,1991,8:168-172
93 KrumbeinW E.Calcification by bacteria and algae[C]∥Trudinger P A,Swaine D J.Biogeochemical Cycling of Mineral-Forming Elements.Amsterdam:Elsevier,l979:47-67
94 Krumbein W E.Photolithottophic and chemoorganotrophic activity ofbacteria and algae as related to beachrock formation and degradation (Gulf of Aqaba,Sinai)[J].eomicrobiological Journal,1979,1: 139-203
95 Malone Ph G,Towe KM.Microbial carbonate and phosphate precipitates from seawater cultures[J].arine Geology,1970,9:301-309
96 WattW D,Hayes F R.Tracer study of the phosphorus cycle in sea water[J].imnological Oceanography,1963,8:276-285
97 Krumbein W E,Cohen Y,Shilo M。1977,Solar Lake(Sinai)4, Stromatolitic cyanobacterialmats[J].imnogical Oceanography,22: 635-656
98 Friedman G M,Sneh A,Owen RW.The Ras Muhammad Pool:implications for the Gavish Sabkha[C]∥Friedman G M,Krumbein W E.Hypersaline Ecosystems:The Gavish Sabkha.Berlin:Springer, 1985,218-237
99 Folk R L.Carbonate petrography in the post-Sorbian age[C]∥Ginsburg R N.Evolving Concepts in Sedimentary.Baltimore:Johns Hopkins University Study Geology,1973,118-158
100 Soudry D,CHhampetier Y.Microbial processed in the Negev phosphorites(southern Israel)[J].edimentology,1983:30,411-423
101 Dahanayake K,Krumbein W E.Ultrastructure of a microbial mat generated phosphorite[J].ineral Deposita,l985,20:260-265
102 Dahanayake K,Krumbein W E.Microbial structure in oolitic iron formations[J].ineral.Deposita,1986,21:85-94
103 Decho AW,Visscher PT,Reid R P.Production and cycling ofnaturalmicrobial exopolymers(EPS)within amarine stromatolite[J].alaeogeography,Palaeoclimatology,Palaeoecology,2005,219: 71-86
104 Van Lith Y,Warthmann R,Vasconcelos C,et al.Microbial fossilization in carbonate sediments:A result of the bacterial surface involvement in carbonate precipitation[J].edimentology,2003,50: 237-245
Brief Introduction on New Advances on the Origin of Ooids
MEIMing-xiang
(School of Earth Sciences and Resources,China University of Geosciences,Beijing 100083)
The origin of ooids represents a sedimentogical problem like an enigma.On the basis of experiment studies,Brehm et al。(2006)concluded that the ooid can be analogical to the stromatolite,i.e.the ooid is the product resulted from the activities of a special sphericalmicrobialmat.Recently,studies on modern ooids in Bahamian Archipelago by Duguid et al。(2010)indicate thatmicrobes do not play a role in ooid formation;thus,a new chemical model for the ooid formation has been proposed,i.e.whereby a veneer of ACC precipitates on an ooid while it is at the sedimentwater interface(the active phase)and this veneer of ACC later recrystallizes to aragonite needles and a new cortex layer is formed(the stationary phase).wo different views represent the new advances on studies of the ooid origin.Whereas,the particularmicroscopic fabrics for both the giant ooids of the lower Triassic in south China and the ooids of the Cambrian in north China providemore supporting evidence for themicrobial origin of the ooid formation.
microbial action;chemical precipitation of ACC;ooid origin;researching advances
梅冥相 男 1965年出生 教授 博士研究生导师 沉积学和地层学 E-mail:meimingxiang@263.et
P512.2
A
1000-0550(2012)01-0020-13
①国家自然科学基金项目(批准号:49802012,40472065)资助。
2011-02-04;收修改稿日期:2011-04-25
