基于贝叶斯估计的初级视皮层光栅朝向编码研究
2012-12-03师黎,王艺
师 黎,王 艺
(郑州大学电气工程学院,河南郑州450001)
0 引言
视觉是动物接受外界信息的主要途径,哺乳动物有80%以上的信息是通过视觉系统得到的.朝向是构成图像的基本单元,对朝向进行编码是动物初级视觉皮层(V1区)神经元感受野的基本特性之一[1].研究V1区神经元的朝向编码是视神经科学领域的一个重要研究问题,对于认知动物视觉系统信息处理机制具有重要作用,也是目前国内外的研究热点.
锋电位(spike)是神经元对朝向编码的主要手段,平均发放率和发放时间间隔(ISI)是spike传递视觉刺激响应的两个重要指标[2].目前,spike平均发放率是国内外研究V1区朝向编码最常用的方式[3-9].Adrian最先提出了利用固定时间窗中神经元spike发放数目编码外界刺激信息的方式[3];Barlett和Ralph等人根据平均发放率研究了猫V1区神经元不同生理时期的朝向调谐特性的区别[4];Tan和Brown等人亦是通过V1区神经元的平均发放率研究了大鼠和猫V1区对光栅朝向的编码[5];Matteo等根据平均发放率研究了成年猫V1区简单细胞与复杂细胞对不同朝向的编码[6];Sergej等发现发放率可以在一定程度上反映神经元朝向编码特性[7].近年来,国内外神经生物学研究表明神经元的ISI特性在神经元对视觉刺激信息传递的过程中也起着重要作用.Abeles在研究运动皮层神经元响应时候发现ISI特性与外界刺激的时间有关;Pissadaki和Sidiropoulou等人发现响应中的ISI特性能够准确地对刺激信号进行时间紧密度和空间聚散度分类[8];Reich等发现了猴V1区ISI携带与视觉刺激时间相关的信息[9].但是,V1区神经元 ISI如何编码朝向还是一个需要深入研究的问题.
因此,笔者基于贝叶斯估计理论提出了一种综合大鼠V1区神经元发放率和ISI特性的朝向复合编码方法.分析结果显示:笔者提出的复合编码方法对朝向编码的正确率明显高于仅用发放率编码的结果.这也证明了大鼠V1区神经元的发放时间间隔与发放率共同编码了与光栅朝向相关的刺激信息.
1 材料部分
1.1 准备及手术
健康Long Evens大鼠,雄性,河南省实验动物中心提供,体重200~300 g,外眼及检眼镜检查无异常.按体重4 ml/kg腹腔注射体积分数为10%的水合氯醛对大鼠进行麻醉.根据大鼠脑组织谱图,在V1区开大小为5 mm×2.5 mm的方块区域,将材质为铂铱合金的MicroProbe电极植入,连接多通道生理信号采集系统,以观察V1区神经元放电信号.用于视觉刺激的液晶显示器置于大鼠眼睛前20 cm处.
1.2 光栅刺激
刺激图案采用不同朝向的全屏正弦光栅图像,由MATLAB软件生成.如图1所示.

图1 不同朝向的全屏光栅Fig.1 The gratings with different orientations
光栅的朝向范围在0°~315°之间,以45°为间隔,每隔1 s随机转换一次朝向,每个朝向光栅持续显示相同的时间.一组试验中各个朝向的光栅均显示一次.光栅刺激的时间频率为4 Hz,空间频率为0.2 cpd,对比度为0.8,在进行刺激的过程中,这些参数均固定不变.
1.3 动作电位信号采集
数据采集应用Cerebus-128多通道微电极阵列信号采集系统,共有16个有效通道.原始数据带通滤波(250~5 000 Hz)得到高通连续信号,采用阈值检测的方法提取动作电位.此外,插入大鼠V1区中的一根电极往往引入了若干个神经元的放电信号.因此,为了能够更加准确地计算单个神经元细胞的发放率,还需要对各个通道中的动作电位进行分类.笔者采用PCA降维和K-均值聚类相结合的分类方法[10],以分类后的信号为依据,对光栅朝向进行推断.
2 综合Spike发放率和时间间隔的光栅朝向复合编码方法
笔者依据贝叶斯估计理论研究神经元对光栅朝向的编码.首先,提取神经元spike的发放率和时间间隔两种响应特征;然后,依据多峰高斯模型拟合各个朝向的概率分布密度函数,根据概率密度函数采用综合发放率和时间间隔的复合编码方法对光栅朝向进行编码.
2.1 神经元发放率和时间间隔的提取
提取发放率,步骤如下:
1)设定生理时间窗:Δt=ti+1-ti,通常设为200 ms;
2)计算生理时间窗[ti,ti+1)内神经元的发放个数为Ri;
3)计算生理时间窗[ti,ti+1)内发放率为ri=Ri/Δt,由于实验中刺激持续的时间为1 s,故在一次刺激期间,可以计算出5个发放率,即i=0,1,2,3,4;
5)重复进行实验,同一朝向的光栅si会重复刺激n次,按照上述步骤统计每一次si出现时的平均发放率,记为
统计发放时间间隔信息,步骤如下:
1)假设τi为神经元的发放时刻,计算在光栅刺激期间相邻神经元发放的时间间隔Δτ=τi+1-;
2)统计一次光栅si刺激下神经元所有的发放时间间隔集合
3)计算时间间隔集合的标准差,记为ti;
4)重复进行实验,同一朝向的光栅si会重复刺激n次,按照上述步骤统计每一次si出现时的时间间隔标准差,记为(ti1,ti2,…,tin).
2.2 光栅朝向的编码
根据贝叶斯估计理论可知[11-12]:
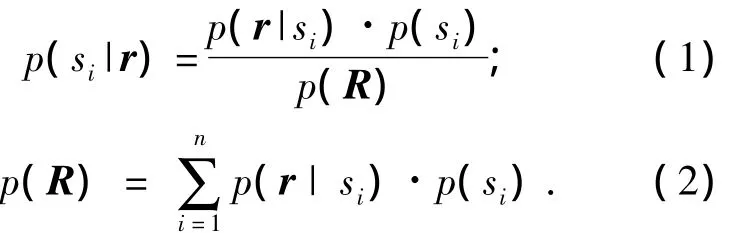
根据(1)和(2)可得:

式中:si为刺激特征,即光栅朝向;r为神经元响应;p(si)为实验中朝向si出现的概率,也称为先验概率;p(r|si)为刺激是si时神经元响应为r的条件概率;p(si|r)为观察到神经元响应为r时刺激为 si概率,也称为后验概率[13].式(3)为依据神经元响应编码光栅朝向的主要依据.笔者判定后验概率值最大时的si为依据r编码得到的光栅朝向,若与实验室刺激的朝向一致,则编码结果正确,否则是错误的.
笔者以神经元的发放率和时间间隔响应作为编码光栅朝向的指标,分别以 r-,t来表示,依据公式(3)可知:
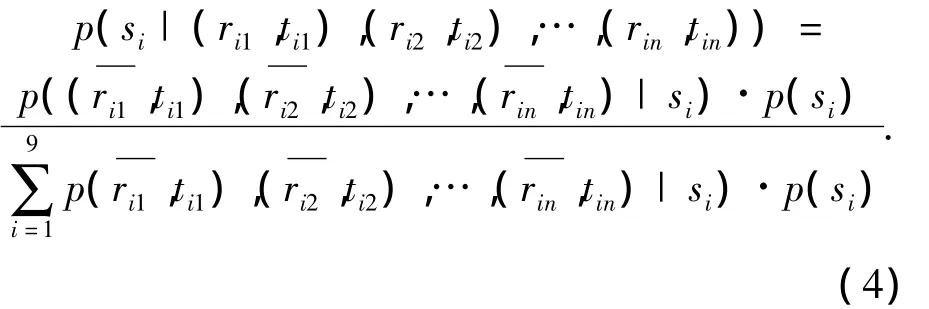
实验中的各朝向光栅是等概率随机出现的,故p(si)=0.125;在实验中发现,即使重复相同的刺激,同一神经元的发放率响应和依然是不同的.笔者考虑每一次光栅si刺激下神经元的响应,假定它们是相互独立的,对这些数据进行高斯拟合,可以得到如下结果:
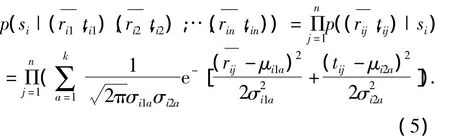
根据式(4)、(5)便可计算出后验概率值.
3 结果与分析
实验数据来源于2011年3月—2011年7月,郑州大学实验平台使用微电极阵列采集的LE大鼠V1区神经元动作电位序列,随机选取10只大鼠,对150个神经元的响应数据进行了分析.
首先,笔者对神经元的朝向选择特性进行了分析[14],限于篇幅,绘制了其中6个具有代表性的神经元朝向调谐坐标示意图,如图2所示.性是不同的.22#、23#、28#对其中少数朝向的选择性强度明显优于其它朝向,而26#、25#、30#对各朝向的选择性强度较为平均.这些神经元朝向选择特性的差异能够体现V1区大部分神经元朝向选择特性的区别.依据神经元的朝向选择性强度分为两种:一种神经元对少数朝向的选择性强度明显优于其它朝向,另一种神经元对各个朝向的选择性强度差别不大.
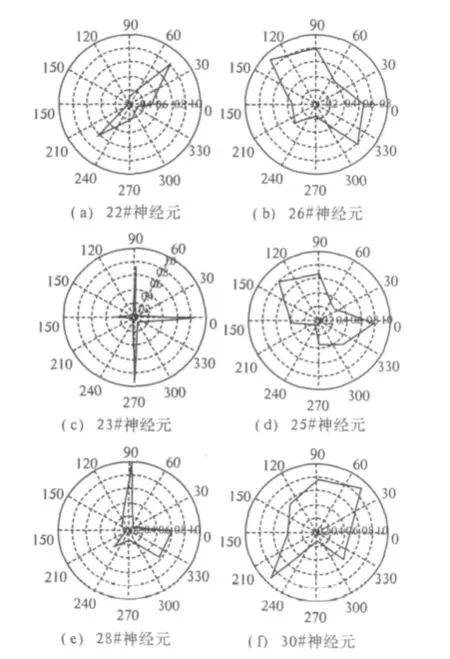
图2 不同神经元朝向调谐曲线的极坐标示意图Fig.2 The orientation tuning curves of different cells
笔者提取了神经元的发放频率与发放时间间隔的响应特性,分别采用复合编码和基于发放率编码的方式对光栅朝向进行编码,并将两种编码结果进行了统计对比分析.以22#、26#为例,对结果进行分析比较,表1统计了依据22#和26#神经元的响应数据,采用发放率和复合编码方法对各朝向编码正确率统计.可以看出,无论是朝向调谐特性较为明显的22#神经元还是朝向调谐特性不明显的26#神经元,复合编码的正确率明显高于发放率编码;22#神经元复合编码正确率比发放率编码结果提高10%左右,26#神经元复合编码正确率比发放率编码结果提高20%左右.证明了发放率与时间间隔共同参与了对朝向信息的编码,而发放率与时间间隔在对朝向进行编码时的相辅相成在26#朝向选择性强度平均的神经元上体现得更为显著.
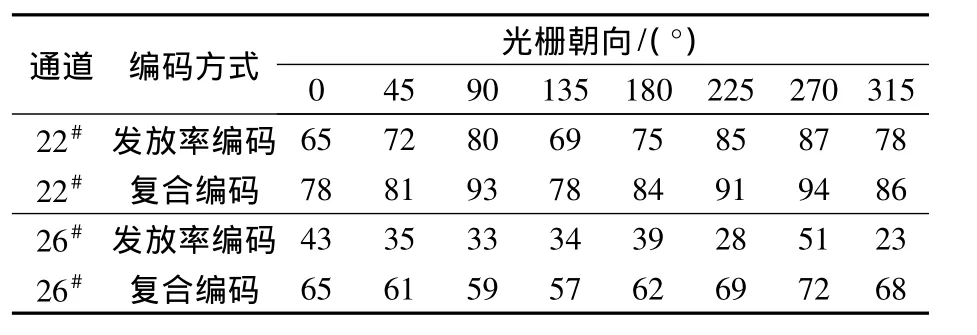
表1 发放率编码和复合编码神经元下各朝向的正确率Tab.1 The accuracy statistic to all orientations estimated between response based on rate encoding and hybrid encoding of neurons selected %
笔者还比较了采用复合编码方式对两种神经元进行编码的正确率,同样以22#和26#为例,结果如表2所示.
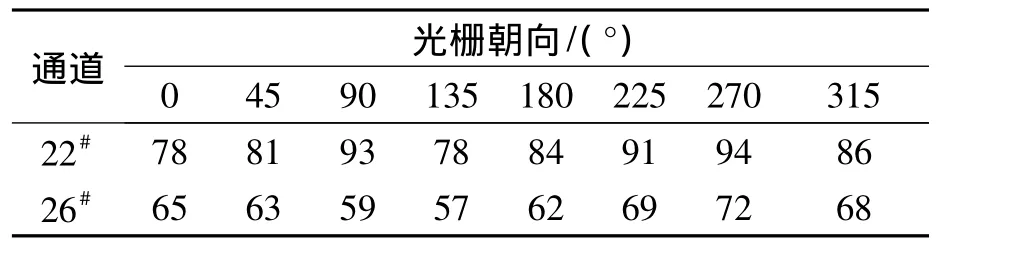
表2 复合编码下22#和26#神经元进行朝向编码的正确率Tab.2 The accuracy statistic to all orientations based on hybrid encoding of the neurons selected %
可以看出,对朝向调谐特性较为明显的22#神经元而言,复合编码的正确率较高,几乎都在75%以上,而对各朝向选择性强度较为平均的26#神经元来说,正确率相对较低.
为了验证复合编码方式的个体适应性,笔者随机选取10只大鼠进行实验,采用朝向调谐特性显著的神经元的响应数据进行复合编码,并对正确率进行统计,如图3所示.

图3 对10只大鼠进行朝向编码的正确率统计Fig.3 The accuracy statistic oforientations estimated of ten rats
从图3可以看出,对每一只大鼠而言,正确率都在78%以上,可见复合编码方式是较为稳定的,个体差异不明显.
4 结论
基于贝叶斯估计理论采用综合发放率和时间间隔编码特性的复合编码方式对光栅朝向进行了编码,并与单独以神经元发放率为响应指标的编码结果进行了统计分析.
(1)神经元的朝向调谐特性较为复杂,表现出较强的不一致性[15],基本上可以分为两类,一类对某些朝向表现出较强的响应特性,而另一类对所有朝向表现出较为平均的响应特性.
(2)复合编码方式的正确率比基于发放率编码方式的正确率高,证明发放率和发放时间间隔共同参与了对朝向的编码,而且正确率的提高对朝向选择特性不明显的神经元表现得更为显著.
(3)比较两类神经元发现,对朝向选择特性较为明显的神经元复合编码的正确率要高于朝向选择特性不明显神经元;复合编码方式较为稳定,个体差异性不明显.
因此,所提出的复合编码方法能更好解析大鼠V1区神经元的信息处理机制,这对研究神经元对外界刺激信息特征的编码具有重要的意义.
[1] SHUSHRUTH S,MANGAPATHY P,ICHIDA J,et al.Strong recurrent networks compute the orientation tuning of surround modulation in the primate primary visual cortex[J].Neuronscience,2012,32(1):308-321.
[2] DUCHATEAU J,ENOKA R.Human motor unit recordings:origins and insight into the integrated motor system[J].Brian Research,2011,1409:42-61.
[3] DEGGER M,HELIASM,BOUCSEIN C,et al.Statistical properties of superimposed stationary spike trains[J].Computional Neuronscience,2011,32(3):443-463.
[4] BARTLETT D,MOORE I,RALPH D,et al.Development oforientation tuning in simple cells of primary visual cortex[J].Neurophysiology,2012,107(9):2506-2516.
[5] TAN A,BROWN B,SCHOOL B,et al.Orientation selectivity of synaptic input to neurons in mouse and cat primary visual cortex[J].Neuroscienc,2011,31(34):12339-12350.
[6] MATTEOC,DAVID F.Membrane potential and firing rate in cat primary visual cortex [J].Neuroscience,2000,20(1):470-484.
[7] SERGEJ V,YVES S,RAYMOND D,et al.Receptive field properties of single neurons in rat primary visual cortex [J].Neurophysiology,1999,82:301-311.
[8] PISSADAKI E,SIDIROPOULOU K,RECZKO M,et al.Encoding of spatio-temporal input characteristics by a CA1 pyramidal neuron model[J].PLoSComputional Biology,2010,6(12):1-19.
[9] REICH D,MECHLER F,PURPURA K,et al.Inter spike intervals,receptive fields,and information encoding in primary visual cortex [J].Neuronscience,2000,20(5):1964-1974.
[10]杨振兴.大鼠初级视觉皮层V1区视觉刺激的响应信号分析[D].郑州:郑州大学电气工程学院,2011.
[11] JEFFEREY M.,ROOZBEH K.Probabilistic population codes for bayesian decision making[J].Neuron,2008,60:1142-1152.
[12] ROGRIGO Q,STEFANO P.Extracting information from neuronal populations:information theory and decoding approaches[J].Neuronscience,2009,29(10):173-185.
[13] DARIO L,MICHEL J,ROBERT S,et al.Dynamics oforientation tuning in Macaque V1:the role of global and tuned suppression [J].Neurophysiology,2003,90:342-352.
[14] ALEXANDER P,PETER D,RICHARD Z,et al.Information processing with population codes[J].Neuronscience,2000,20(1):125-132.
[15] JAMESA,WILLIAME.Spatial frequency and orientation tuning dynamics in area V1 [J].PANS,2005,99(3):1645-1650.
