力竭运动过程中大鼠纹状体神经元局部场电活动的动态研究
2012-11-17乔德才李许贞刘晓莉胡琰茹
乔德才 李许贞 刘晓莉 胡琰茹
北京师范大学体育与运动学院(北京 100875)
本实验室从2001年开始关注纹状体对运动性疲劳(exercise-induced fatigue)的中枢调控作用及其机制的研究,通过采用玻璃微电极细胞外电生理记录技术,麻醉状态下在体观察发现,运动疲劳大鼠纹状体神经元自发高频放电比例增加,出现爆发式放电活动特征[1],初步证实纹状体神经元参与了运动疲劳的中枢调控,也是引起运动疲劳的重要中枢脑区之一。但由于实验存在一定的滞后性,上述研究结果只能反映大鼠疲劳时神经元电活动与安静状态的差异。事实上,大鼠在运动过程中的行为能力具有明显的阶段性特征[2,3],其行为能力改变主要受运动中枢神经元功能的控制与调节[4,5]。 为此,本研究采用金属记录电极手术植入技术,实现对大鼠一次性力竭运动过程中纹状体神经元电活动的实时、动态观察,探讨安静、运动、疲劳、恢复等不同状态下纹状体神经元电活动变化特征与运动能力的关系,为揭示纹状体在运动疲劳形成、恢复过程中的调控规律提供参考。
1 材料与方法
1.1 实验动物
健康雄性 Wistar大鼠 12 只,体重(250±10)g,购自北京维通利华实验动物技术有限公司。动物生产许可证号:SCXK(京)2002-2003。常规分笼饲养,自由饮水进食,自然光照,动物房内温度(20±3)℃,相对湿度为40%~60%。本实验在北京师范大学运动生理学实验室完成。
1.2 金属电极植入手术
实验前将大鼠用戊巴比妥钠(50 mg/kg)腹腔麻醉,俯卧固定于脑立体定位仪(SN-3N,日本成茂)上,沿大鼠头顶正中线做一矢状位切口,暴露前、后囟及冠、矢状缝等骨性标志,调整门齿高度,使前囟和后囟处在同一水平面上。依照大鼠脑立体定位图谱[6],在纹状体对应的颅骨部位钻孔,掀去硬脑膜,将金属电极植入左侧纹状体(P:0.2,L:3,H:3.2),并用小螺钉和牙科水泥固定,保证电极不随动物的活动而移动或松动。注射地塞米松注射液缓解术后脑水肿。大鼠恢复4~5天,手术引起的不良反应即可消失,饮食和行为活动恢复正常,开始进行恢复性递增负荷运动训练,并使大鼠逐渐适应测试系统导联状态下的跑台运动。待大鼠能以20 m/min的速度跑30 min,并未见不良反应,即可进行正式实验。
1.3 大鼠力竭运动方案
力竭运动采用本实验室根据Bedford方法修改建立的递增负荷跑台运动方案[7]。运动前,先将大鼠置于动物跑台 (DSPT-202,杭州段氏)上安静30 min,随后进行跑台运动。负荷分三级:I级负荷:8.2 m/min,15 min;II级负荷:15 m/min,15 min;III级负荷:20 m/min,运动至力竭。力竭判断标准为:大鼠跑姿由蹬地式变为伏地式,滞留在跑道末端不能继续跑动,且声波和光刺激均不能驱使动物继续维持跑动。
1.4 大鼠力竭运动过程中LFPs信号采集
大鼠开始运动之前,金属电极经导联线与主放大器(Dagan EX4-400,美国)连接,导联线由柔细屏蔽线组成,以尽量减少工频干扰对信号的影响。导联线靠近电极端包含一个JFET前置放大器(headstage),该前置放大器由一块TL082(Texas Instrument,美国)集成电路制成,用于消除运动过程中由于导线运动、缠结造成的信号伪迹。信号采集由多通道记录仪 (PowerLab 8/30,AD Instrument澳大利亚)及自身所带软件Chart5完成,采样频率512 Hz,50 Hz数字陷波,主放大器硬件滤波设置为0.1~100 Hz,增益200。连续动态记录大鼠安静、运动、力竭和恢复期纹状体神经元局部场电位 (local field potentials,LFPs)的电信号(图1),并对大鼠运动能力进行观察。实验完成后,取脑组织制作常规冰冻切片,并进行Nissl染色,对照大鼠脑立体定位图谱鉴定金属电极尖端所在位置(图2),金属电极未准确插入纹状体的数据将被删除。
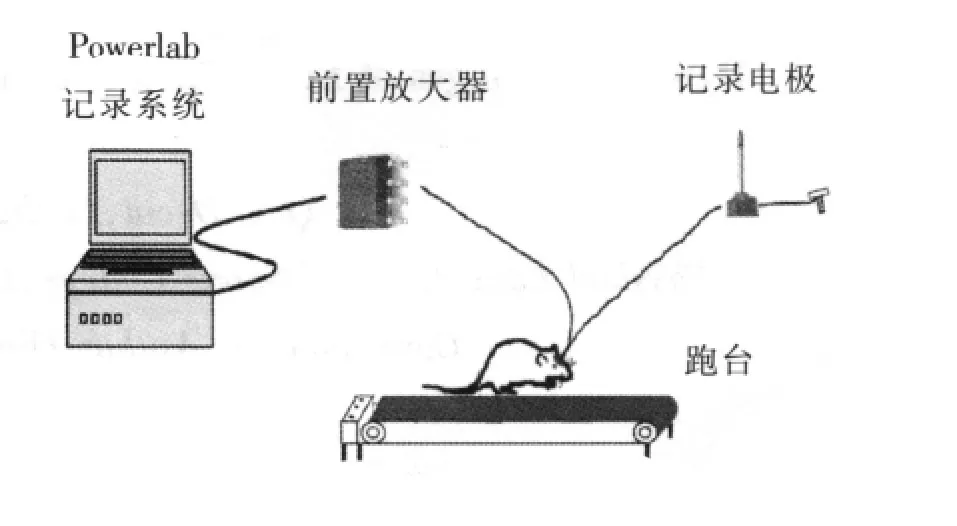
图1 力竭运动过程中纹状体LFPs动态记录模式图
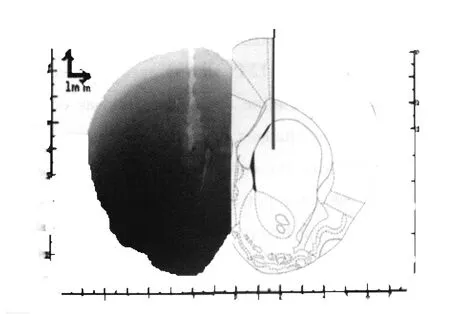
图2 纹状体局部电极尖端定位脑片图
1.5 数据统计与分析
使用 Chart5 及 NeuroExplorer4(Plexon,美国)离线分析软件,快速傅立叶转换 (Fast Fourier Transform,FFT)welch 法(FFT size为 512),对原始 LFPs信号进行各种参数的统计与分析。从力竭运动过程中每一阶段选取4个30s的原始信号进行频率谱分析。根据脑电活动特征划分的方法,将LFPs划分为5个频段 (δ 波:0.8~3.9 Hz,θ 波:4~7.9 Hz,α 波:8~12.9 Hz,β 波:13~30 Hz 及 γ 波:30 Hz以上)[8], 分别计算不同频段的功率谱密度值(power spectrum density,PSD)。力竭运动过程中每15min取4个30s LFPs电信号进行功率谱重心频率(gravity frequency,fg)的计算, 其中:fg为重心频率;f1到 f2为频率范围;P(f)为信号的功率谱;f为频率值[5],计算公式如下:
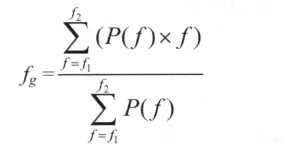
2 结果
2.1 力竭运动过程不同阶段划分及大鼠运动能力观察
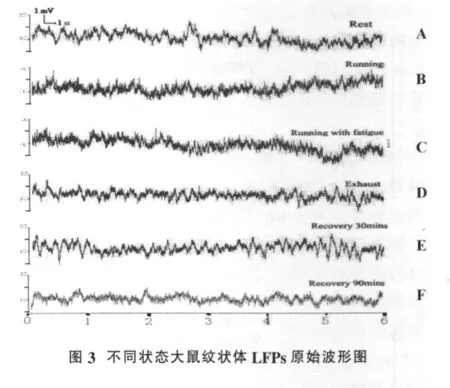
12只大鼠中,实际记录到有效信号的8只,其余大鼠均不纳入数据统计。运动能力观察发现:运动开始后,大鼠进行递增负荷自主运动;持续一段时间后,大鼠的自主运动能力明显下降;此时给予声、光、电等外部轻微刺激后,大鼠仍可维持原有速度继续运动直至力竭。由于不同大鼠运动至力竭的时间个体差异较大(163.7±29.32)min,为便于观察大鼠在不同运动状态LFPs的动态变化规律,根据大鼠行为特征将整个运动过程划分为以下6个阶段:A:安静状态;B:自主运动状态(0~30 min,大鼠在自我驱动下能够维持预定强度运动);C:疲劳运动状态(力竭前60 min~力竭,通过轻度驱赶刺激大鼠能够维持预定强度运动);D:力竭状态(通过驱赶刺激也无法维持预定强度运动);E:恢复30 min(力竭后恢复30 min);F:力竭后恢复 90 min(力竭后恢复 90 min)。忽略自主运动状态与疲劳运动状态之间的时间点,以保证检测样本统计处理的一致性。不同状态下LFPs原始波形如图3所示。
2.2 力竭运动过程中纹状体LFPs频率谱特征变化
力竭运动过程中纹状体LFPs频率谱变化特征如图4所示。从图4中可以看出,安静状态下大鼠纹状体LFPs放电振幅较高,自主运动状态下放电幅度明显降低(图4-A);安静、力竭和恢复30 min纹状体LFPs频率谱有明显差异,力竭状态放电幅度明显低于安静状态和恢复期(图4-B)。
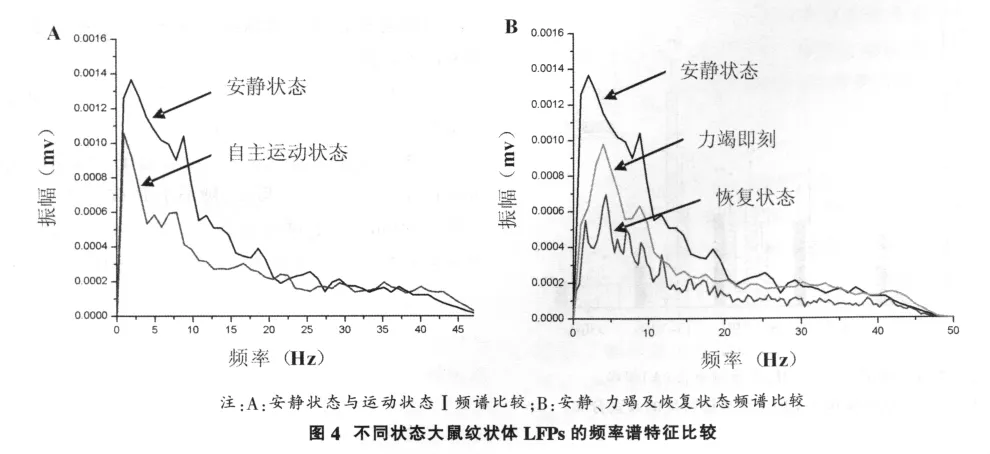
2.3 力竭运动过程中纹状体LFPs功率谱特征变化
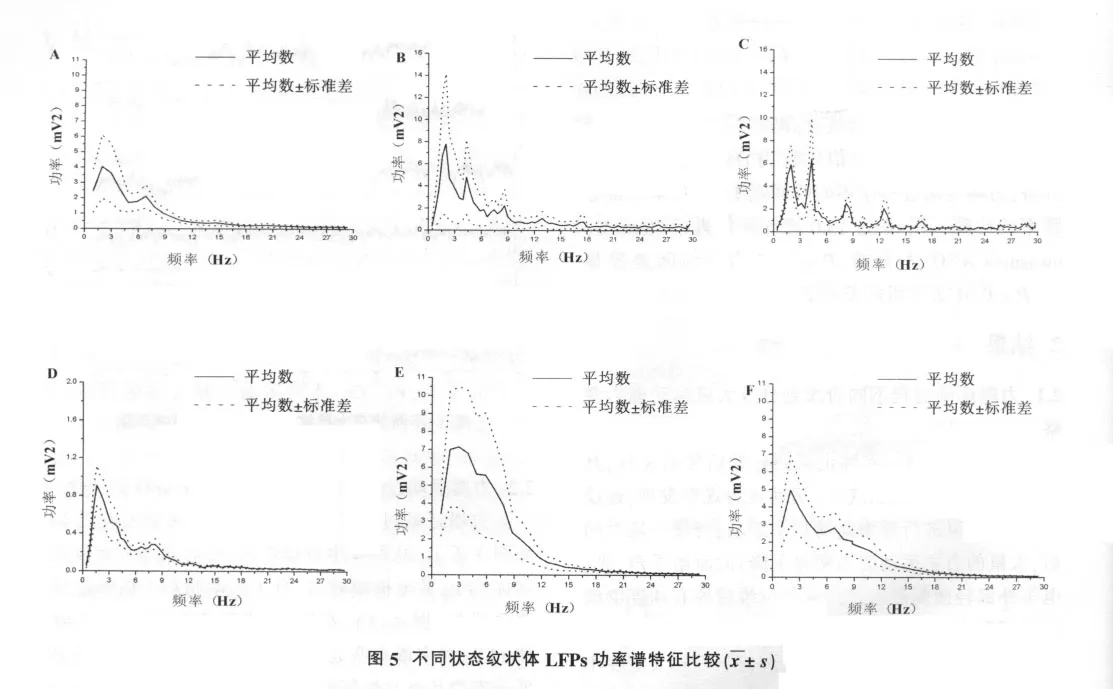
力竭运动过程中纹状体LFPs功率谱变化特征如图5所示。从图5中可以看出,大鼠纹状体LFP功率谱分布在运动状态与非运动状态有明显差异,主要表现在运动状态总功率出现多峰值,特别是疲劳运动状态;在安静、自主运动、力竭及其后的恢复期内,大鼠纹状体LFP放电频率主要集中在<10Hz范围内,以慢波δ和θ波为主;而疲劳运动状态下大鼠纹状体LFP放电频率主要集中在<15Hz范围内,α波增多。
以安静状态下各频段的PSD值为基值,求其它阶段各频段与安静状态的比值,并对比值进行统计,以消除不同大鼠个体差异带来的影响(图6)。从图6可以看出,与安静状态相比,自主运动和疲劳运动状态纹状体LFPs各波段PSD显著升高 (P<0.05,P<0.01);力竭时各波段PSD明显下降至安静水平;恢复30 min和90 min,δ波、θ波、α波PSD仍显著高于安静状态(P<0.05),而β波、γ波PSD则均未出现显著变化(P>0.05)。
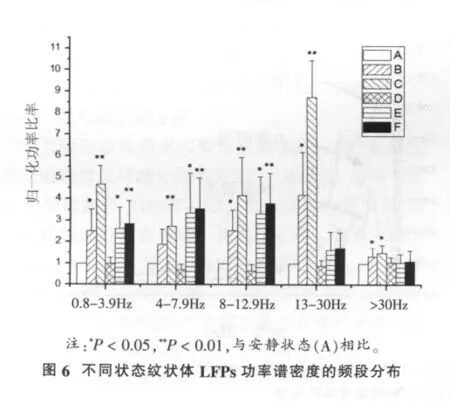
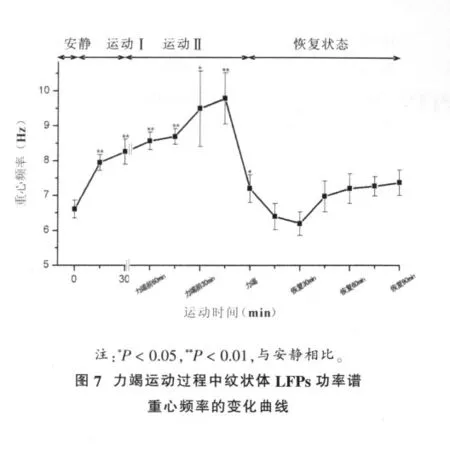
2.4 力竭运动过程中大鼠纹状体LFPs功率谱重心频率的变化曲线
力竭运动过程中大鼠纹状体LFPs功率谱重心频率的变化,结果如图7所示。从图7中可以看出,运动开始后,fg持续显著升高 (P<0.05,P<0.01),力竭前15 min升至最高点,随后fg持续下降,力竭后恢复30 min时fg降至最低点,之后虽略有回升,但与安静水平相比无显著性差异(P>0.05)。
3 讨论
基底神经节与运动控制有密切联系[9]。通过直接通路(皮层―纹状体―苍白球内侧部/黑质网状部―丘脑―皮层)、间接通路(皮层―纹状体―苍白球外侧部―丘脑底核―苍白球内侧部/黑质网状部―丘脑―皮层)和超直接通路(皮层―丘脑底核―苍白球内侧部/黑质网状部―丘脑―皮层)与皮层运动区发生联系[10,11]。直接通路可对运动皮层产生兴奋作用,间接和超直接通路可对运动皮层产生抑制作用。纹状体是基底神经节中信息传导的主要核团之一,它接受来自皮层的大量神经投射,直接或间接参与随意运动的程序编制与执行。同时,在调节运动方向、顺序、速度、幅度以及运动可塑性等方面都具有重要作用[12]。目前,对纹状体神经元活动机制的研究主要采用电生理学研究方法,它可以在不伤及组织的条件下观察神经元活动的瞬间变化。记录LFPs是神经电生理学研究的重要手段之一,主要分析指标有放电频率及频率谱、功率谱密度及功率谱重心频率等。低频、高幅的神经元放电活动一般代表中枢神经系统兴奋性降低,抑制性增强;而高频、低幅则反映中枢神经系统兴奋性增强,抑制性减弱。功率谱重心频率曲线不仅可以较好地反映功率谱密度的分布情况,还反映不同条件下整个功率谱的迁移情况,它与脑功能状态存在非常密切的关系[8]。清醒安静时,正常脑电的高、低频放电功率相对平衡;脑疲劳时,皮层抑制程度增加,脑电高频成分减少,低频成分增加,整个功率谱重心频率向低频迁移,反之,则向高频迁移。
本研究结果发现,一次性力竭运动过程中,大鼠纹状体神经元LFPs活动的动态变化特征主要表现为:安静状态下放电频率较低、幅度较高;运动过程中放电频率逐渐升高、幅度逐渐降低;力竭后逐渐恢复至安静时水平。功率谱重心频率动态变化规律为:运动开始,纹状体LFPs活动的功率谱重心频率由低频向高频逐渐迁移,力竭前达到峰值,随后逐渐向低频迁移,力竭后及恢复初期降至最低点。由此可以认为:当纹状体接受来自大脑皮层的运动指令后,通过直接通路向皮层发放兴奋性冲动,以发起动作并维持预定的运动强度;当运动时间持久并出现疲劳时,纹状体通过间接通路向皮层发放抑制性冲动,使皮层兴奋性减弱,导致运动能力降低。纹状体在力竭运动的不同阶段可分别通过直接通路和间接通路对运动行为能力进行调节。
神经元间的信息传递主要依赖神经递质实现[12]。纹状体可接受来自皮层的谷氨酸(Glu)能兴奋性神经纤维投射,再发出γ-氨基丁酸(GABA)能神经纤维,经直接或间接通路到达苍白球内侧部/黑质网状部,经丘脑返回皮层[13]。本实验室前期采用微透析和高效液相色谱联用等技术发现,大鼠一次性力竭运动过程中在自主运动期,纹状体内兴奋性神经递质Glu的作用占优势,神经元以兴奋性功能活动为主,通过激活直接通路,释放更多的GABA抑制苍白球内侧部/黑质网状部的活动,对丘脑GABA能神经元去抑制,对运动起兴奋作用;而在疲劳运动期和力竭期,纹状体内抑制性神经递质GABA活动占优势,GABA能纤维主要通过间接通路,使丘脑底核Glu能神经元去抑制,进而引起苍白球内侧部/黑质网状部的兴奋性增强,抑制丘脑Glu神经元活动,最终对运动起抑制作用[14,15]。 因此,可以推测,一次性力竭运动过程中大鼠在自主运动期纹状体主要是以Glu能神经元功能活动占优势,而在运动疲劳后期至力竭期,则是以GABA能神经元的功能活动占优势。
4 总结
一次性力竭运动过程中大鼠纹状体神经元LFPs活动呈现动态变化规律,主要表现为:与安静状态相比,运动过程中放电频率逐渐升高、幅度逐渐降低,力竭后逐渐降低至安静水平。纹状体LFPs电活动的改变与运动疲劳有关。可以认为,在运动初期纹状体主要是通过直接通路参与皮层运动的调控,而在运动后期,则主要通过间接通路发挥作用。提示:两条通路的平衡失调是导致大鼠运动能力下降的重要原因之一。
[1]乔德才,侯莉娟,何德富,等.运动疲劳对大鼠新纹状体神经元电活动的影响.中国运动医学杂志,2005,24(6):676-680.
[2]刘晓莉,罗勇,乔德才.大鼠一次性力竭跑台运动模型的建立与动态评价.中国实验动物学报,2012,20 (3):25-28.
[3]Secher NH,Quistorff B,Dalsgaard MK.The muscles work,but the brain gets tired.Ugeskr Laeger,2006,168 (51):4503-4506.
[4]Chaudhuri A,Behan PO.Fatigue and basal ganglia.J Neurol Sci,2000,179(1-2):34-42.
[5]王大磊,刘晓莉,乔德才.大鼠力竭运动过程中丘脑底核和皮层神经元电活动的变化.中国应用生理学杂志,2011,27(4):427-431.
[6]Paxinos G,Watson C.The rat brain in stereotaxic coordinates.San Diego:Academic Press,1997,22.
[7]杨东升,乔德才,刘晓莉.力竭运动过程中大鼠纹状体葡萄糖/乳酸代谢的实时观察.中国运动医学杂志,2009,28(4):384-387.
[8]Klimesch W.EEG alpha and theta oscillations reflect cognitive and memory performance:a review and analysis.Brain Res Rev,1999,29(2-3):169-195.
[9]Groenewegen HJ.The basal ganglia and motor control.Neural Plast,2003,10(1-2):107-120.
[10]Middleton FA,Strick PL.Basal ganglia and cerebellar loops:motor and cognitive circuits.Brain Res,2000,31:236-250.
[11]Nambu A,Tokuno H,Takada M.Functional significance of the cortico-subthalamo-pallidal “hyperdirect” pathway.Neurosci Res,2002,43(2):111-117.
[12]Meeusen R,Watson P,Hasegawa H,et al.Brain neurotransmitters in fatigue and overtraining.Appl Physiol Nutr Metab,2007,32(5):857-864.
[13]侯莉娟,刘宏强,乔德才.纹状体神经细胞分型及其功能构建.中国临床康复,2004,8(10):1928-1929.
[14]乔德才,李许贞,杨东升,等.力竭运动前后大鼠纹状体谷氨酸和γ-氨基丁酸水平的动态观察.中国运动医学杂志,2011,30(10):921-925,971.
[15]王大磊,刘晓莉,乔德才.力竭运动大鼠丘脑底核mGluR5/GABA-ARa1表达及MPEP干预对皮层运动区兴奋性的影响.中国运动医学杂志,2010,29 (5):542-546.
