抗吡虫啉、高效氯氰菊酯棉蚜品系的室内筛选及适合度研究
2012-11-10孙磊杨德松
孙磊,杨德松
(石河子大学农学院,石河子832003)
棉蚜(Aphis gossypii Glover)属同翅目蚜科蚜属,其年发生代数多,繁殖力强,是棉花的重要害虫之一[1]。1991、1994和2004年,棉蚜在新疆北疆各地暴发,损失率达10%~30%[2-3]。由于棉蚜世代历期短、发生量大,并且长期大量使用化学农药,加之未能科学用药,从而导致棉蚜抗药性的快速发展,其抗性程度高,抗性背景复杂,这给棉花生产造成了很大的困难[4]。虽然新疆棉花采用了膜下滴灌的栽培方式,但并未影响棉蚜的发生[5],因而化学防治依然是防治棉蚜的首选措施。关于蚜虫抗性品种室内选育的报道较多,帅霞等[6]用高效氯氰菊酯对桃蚜进行抗性选育,筛选3代后相对抗性倍数为13.00,10代后为49.86倍。陈亮[7]用吡虫啉对桃蚜敏感品系在室内进行了抗性选育,测定了桃蚜抗吡虫啉品系对9种常用杀虫剂的交互抗性。杨焕青等[8]在室内用吡虫啉对敏感种群棉蚜连续汰选27代,对吡虫啉抗性达24.38倍。杀虫剂长期使用势必产生抗药性,吡虫啉属于新型烟碱类杀虫剂,在新疆棉区是防治棉蚜首选药剂。
为延长现有农药在新疆棉区的使用寿命,有效控制棉蚜,阐明新疆棉区棉蚜种群对杀虫剂的抗性现状,揭示棉蚜的抗药性机理,本研究利用生命表研究棉蚜的抗性种群稳定性,旨在为合理制定棉蚜的抗性治理措施提供依据。
1 材料与方法
1.1 材料
1.1.1供试虫源
棉蚜敏感品系(S)由石河子大学农学院昆虫教研室提供,该种群是室内经连续2年的无毒饲养,期间未曾接触任何化学农药,选取整齐一致的无翅成蚜为标准试虫。棉蚜抗吡虫啉品系(Im R)与棉蚜抗高效氯氰菊酯系(BeR)是由敏感品系在室内经药剂处理12代。轮换使用吡虫啉和高效氯氰菊酯对敏感品系筛选11代获得交互抗性品系(ImBeR)。上述试虫均是在温度(25±1)℃、相对湿度60%±10%、光周期14 L∶10 D的光照培养箱中培养。
1.1.2供试药剂
95.5 %吡虫啉(WP)、95%高效氯氰菊酯(WP)由南京卓创化学有限公司提供。
1.2 方法
1.2.1生物测定方法
测定方法采用叶片浸渍法[9]。将有蚜虫的棉花叶片分别浸入不同浓度药液10 s,取出后让叶片表面水滴自然挥发,反面朝上放置在培养皿中,24 h后检查死蚜数,检查时用毛笔拨动虫体,试虫完全不动视为死亡。若对照死亡率>20%,则当天全部处理作废,若对照死亡率<20%为有效测定。每种药剂设置5个浓度,以丙酮为对照,每浓度处理20头,3个重复。根据死亡机率值和浓度对数值求出毒力回归式(Y=A+BX)、致死中量LD50及诊断量LD99值,并与敏感棉蚜种群进行毒力比较,计算棉蚜的敏感性或抗药性水平[10]。
1.2.2抗性选育数据处理
利用DPS进行数据处理,求出敏感种群与抗性种群的毒力回归方程,求出LD50,95%置信限。抗性倍数等于棉蚜抗性种群LD50与敏感种群LD50的比值抗性指数,计算公式为:

1.2.3抗性适合度研究
1.2.3.1 试验种群生命表的组建
在直径为9 cm的培养皿内用琼脂培养基保湿,将新鲜叶片背面向上铺在培养基上。四周用脱脂棉棉条缠绕,用来保湿并防止棉蚜逃逸。每一叶片接同一天若蚜1头,4-5 d更换1次新鲜棉花叶片,逐日观察记录蜕皮、死亡率,待产蚜后记录日产蚜量,并剔除当日产下蚜虫,直到棉蚜全部死亡。
1.2.3.2 数据处理
参照丁岩钦[11]的方法,组建棉蚜的实验种群生命表,计算生命表各项参数。用DPS和Excell软件对所得数据进行处理。

其中:R0为种群净增殖率,rm为种群内禀增长力,T为世代周期,λ为周限增长率,Dt为种群加倍时间。
1.2.4解毒酶活性测定
酶源蛋白质含量测定参照Bradford[12]的考马斯亮蓝G-250染色法;羧酸酯酶(Car E)活性测定参照Han等[13]的方法;酸性、碱性磷酸酯酶(ACP、ALP)活性测定参照高希武等[14]的方法;谷胱甘肽S-转移酶(GSTs)活性测定参照 Oppenoorth[15]的方法。
1.2.5多功能氧化酶(MFO)活性测定
以对硝基苯酚量为自变量,OD值为因变量作标准曲线。用对硝基苯甲醚(丙酮为溶剂,0.05 mol/L)作底物,在氧和NADPH作电子供体条件下,MFO催化发生氧脱甲基作用生成对硝基苯酚。用盐酸(1 mol/L)终止反应后,先后用氯仿、NaOH溶液(0.5 mol/L)萃取,在400 nm处测 OD值。根据对硝基苯酚标准曲线和酶源蛋白质含量,将OD值换算成比活力[nmol/mg·Pro./30min)],重复3次。
2 结果与分析
2.1 棉蚜对吡虫啉和高效氯氰菊酯的抗性及交互抗性
由于吡虫啉和高效氯氰菊酯的作用机理不同,在抗性选育过程中棉蚜对这两种药剂的抗性发展速度也表现出一定的差异性,吡虫啉汰选敏感棉蚜12代筛选出抗性倍数为7.90的抗性品系,而高效氯氰菊酯汰选12代后获得抗性倍数为10.04的抗性品系(表1)。

表1 吡虫啉对棉蚜抗性筛选动态(2011年,石河子)Tab.1 Resistance development of A.gossypii to Imidacloprid(2011,Shihezi)

表2 棉蚜对高效氯氰菊酯的抗性筛选动态(2011年,石河子)Tab.2 Resistance development of A.gossypii to Beta-cypermethrin(2011,Shihezi)

表3 棉蚜对吡虫啉和高效氯氰菊酯的交互抗性筛选动态(2011年,石河子)Tab.3 Cross resistance development of A.gossypii to Imidacloprid and Beta-cypermethrin(2011,Shihezi)
2.2 敏感品系与抗性品系棉蚜的适合度
不同品系棉蚜的存活率下降趋势基本一致,敏感品系的存活率均高于其他3个抗性品系,表明棉蚜产生抗性以后,其存活率弱于敏感品系(图1)。3种棉蚜品系均能完成其世代,棉蚜的敏感品系的产卵量高于具有抗性的品系表明产生抗性以后产若率有所下降(图2~4)。

图1 棉蚜敏感与抗性品系存活率动态Fig.1 Survival rate between sensitive and resistant strains of A.gossypii
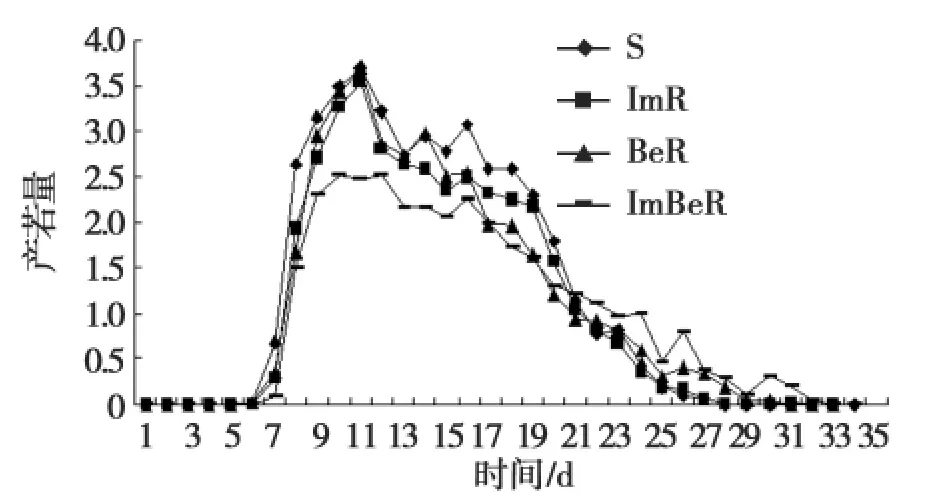
图2 棉蚜敏感品系与抗性品系产仔量日变化Fig.2 Diurnal variation of fertility between sensitive and resistant strains of A.gossypii

图3 不同棉蚜种群世代产若量Fig.3 The amount of generation Nymphs producing by different populations of A.gossypii

图4 棉蚜敏感品系和抗性品系日产若量Fig.4 The amount of diurnal Nymphs producing by different populations of A.gossypii
棉蚜抗性品系和敏感品系的发育速率、世代历期和成蚜寿命等方面均无显著差异(表4、5)。棉蚜敏感品系的生殖力弱于棉蚜的抗性品系(图2)。根据棉蚜的若蚜存活率和净生殖率等参数建立年龄特征生命表(表5)。棉蚜抗性品系平均日产仔量低于敏感品系,成蚜生殖期短于敏感品系,且日产若量低于敏感品系,结果表明抗性品系的生物适合度明显降低。
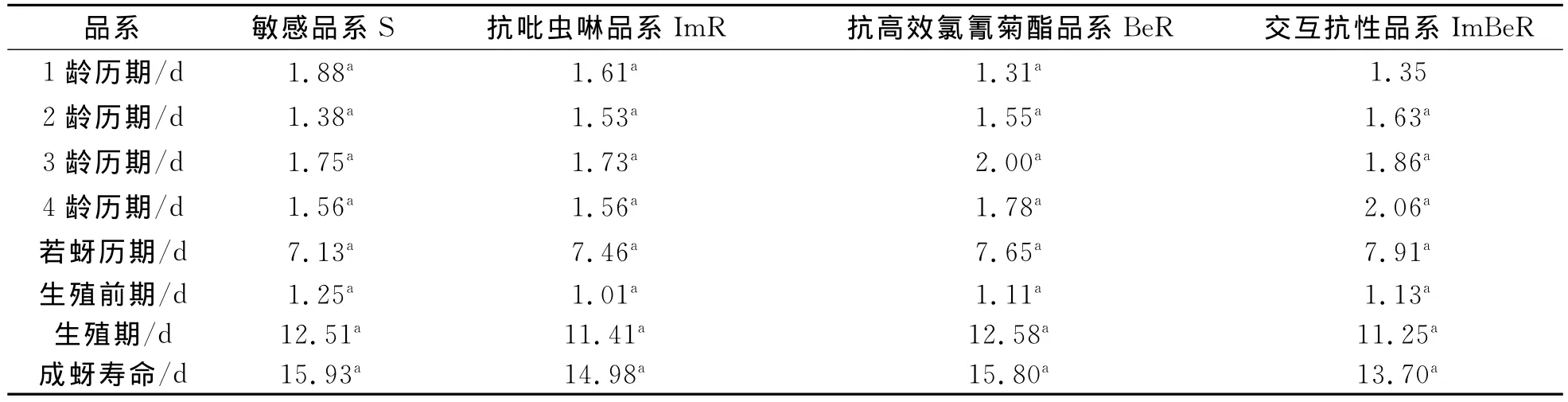
表4 棉蚜敏感品系和抗性品系发育历期比较Tab.4 The comparison of developmental duration between sensitive and resistant strains of A.gossypii

表5 棉蚜的抗性、敏感品系实验种群生命表Tab.5 Life-table of resistant and sensitive populations of A.gossypii
2.3 敏感品系与抗性品系棉蚜解毒酶活性比较
由表6可见,BeR品系棉蚜的羧酸酯酶比活力与敏感品系存在显著差异,高于敏感品系。3个抗性品系棉蚜的酸性、碱性磷酸酯酶、MFO酶与敏感品系无差异性,但谷胱甘肽-S-转移酶与敏感棉蚜相比差异显著,高于敏感棉蚜。说明谷胱甘肽-S-转移酶是解毒吡虫啉和高效氯氰菊酯的主要解毒酶,在解毒吡虫啉和高效氯氰菊酯中起重要作用。

表6 不同棉蚜种群解毒酶比活力Tab.6 Detoxifying enzymes of different populations of A.gossypii
3 讨论
抗性风险评估除了观察抗性的筛选速度外,还需对抗性害虫的适合度进行测定。本研究对吡虫啉和高效氯氰菊酯进行抗性筛选,用吡虫啉选育至12代抗性为7.90(表1),高效氯氰菊酯的抗性选育到12代为10.04(表2),用吡虫啉和高效氯氰菊酯同时选育至11代抗性分别为5.40和6.43(表3)。研究表明,吡虫啉相对于高效氯氰菊酯的抗性发展速度缓慢,说明新疆棉花生产中应用吡虫啉防治棉蚜比高效氯氰菊酯相对安全,但是长期继续使用,可能会产生抗药性。
生物适合度是指一个生物能生存,并且把它的生物学特性传给下一代的相对能力,一般包括繁殖力和生活力等。抗药性往往增加了害虫抗性个体,在药剂选择压力下的存活率,然而抗药性往往导致生物适合度降低,使害虫抗性个体,在没有药剂选择压力时,表现出生存竞争劣势[16]。杨焕青等[8]提出抗吡虫啉品系棉蚜,其产生的若蚜存活率、净生殖率、内禀增长率和周限增长率较敏感品系均有显著降低,相对适合度较小。陈亮[7]研究得出抗吡虫啉桃蚜品系的净生殖率和内禀增长率均低于敏感品系,存在适合度不利性。于金凤等[17]研究发现抗灭多威和抗硫丹的棉蚜种群的繁殖力与敏感品系相比明显降低。吴孔明等[18]发现抗杀灭菊酯高抗品系的若蚜发育历期短于敏感若蚜。刘凤沂等[19]研究报道昆虫抗药性的产生伴有适合度劣势,即抗性个体常表现出发育速率较慢、存活率和生殖力较低。本研究也同样证实3种抗性品系的棉蚜与敏感品系棉蚜相比较在净生殖力、内禀增长率等存在适合度不利性。
羧酸酯酶、磷酸酯酶、谷胱甘肽-S-转移酶和MFOs是昆虫体内重要的解毒酶系,潘文亮等[20]研究抗吡虫啉棉蚜品系中羧酸酯酶和谷胱甘肽-S-转移酶的比活力明显高于敏感品系。帅霞等[6]发现高效氯氰菊酯筛选桃蚜的过程中,LD50与酯酶含量及活性变化之间呈显著正相关,酯酶在桃蚜对高效氯氰菊酯抗药性发展中起着十分重要的作用。吕敏等[21]研究发现谷胱甘肽-S-转移酶是杀虫剂产生代谢抗性的重要酶系,与昆虫抗药性的产生有关。杨焕青[8]测定出抗吡虫啉棉蚜种群的羧酸酯酶和谷胱甘肽S-转移酶比敏感品系高2.73和1.57倍,说明羧酸酯酶和谷胱甘肽-S-转移酶比活力的提高是引起棉蚜对吡虫啉产生抗药性的重要原因之一。慕立义等[22]研究表明敏感品系棉蚜在氰戊菊酯、氧化乐果连续筛选16代后,对氰戊菊酯、氧化乐果产生了抗性及交互抗性,测得3个抗性种群的羧酸酯酶明显高于敏感品系。本研究中发现棉蚜产生抗性与羧酸酯酶和谷胱甘肽-S-转移酶有关。棉蚜ImR、BeR、ImBeR三种抗性品系的谷胱甘肽-S-转移酶与敏感棉蚜相比较显著高于敏感棉蚜;BeR品系的羧酸酯酶显著高于敏感棉蚜,说明这些酶在解毒吡虫啉和高效氯氰菊酯的过程中起作用。因此,生产中建议吡虫啉与一些无交互抗性的药剂进行轮换使用可以阻止或延缓棉蚜抗药性的产生。
[1]崔淑芳,李俊兰.棉蚜的发生、危害与防治[J].中国棉花,2010,37(4):33.
[2]王孝法,孟昭璋,罗教祥.新疆植棉区棉蚜分布为害特点及治理对策[J].中国棉花,1997,24(11):12-13.
[3]冯志超,王永安.新疆北部棉区棉蚜大发生原因及综合防治[J].新疆农业科学,2005,42(4):265-268.
[4]高希武,王政国,郑炳宗,等.六种常用杀虫剂对八种蚜虫的选择性毒性[J].昆虫学报,1990,33(3):274-279.
[5]刘冰,王佩玲,芦屹.不同灌水条件下棉蚜发生的规律,石河子大学学报:自然科学版,2008,26(3):296-298.
[6]帅霞,王进军.桃蚜高效氯氰菊酯抗性选育过程中酯酶含量和活性变化[J].动物学研究,2005,26(1):101-105.
[7]陈亮.桃蚜对吡虫啉抗性的生化机理及其适合度研究[D].杭州:浙江大学,2005.
[8]杨焕青,王开运,王红艳,等.抗吡虫啉棉蚜种群对吡蚜酮等药剂的交互抗性及施药对其生物学特性的影响[J].昆虫学报,2009,52(4):175-182.
[9]孙家隆,慕卫.农药学实验技术与指导[M].北京:化学工业出版社,2009.
[10]唐启义,冯光明.实用统计分析及其DPS数据处理系统[M].北京:科学出版社,2002:603-605.
[11]丁岩钦.昆虫数学生态学[M].北京:科学出版社,1994:153-170.
[12]Bradford M M.A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding[J].Analytical Biochemistry.1976,72:248-254.
[13]Han Z J,Moorses G D,Devonshire A L,et al.Associa-tion between biochemical markers and insecticide resistance in the cotton aphid,Aphis gossypii[J].Pesticide Biochemistry and Physiology,1998,62(3):164-171.
[14]高希武,胡慢华,郑炳宗.应用酶标仪动力学方法监测棉蚜的抗药性[J].昆虫知识,1998,35(1):17-19.
[15]Oppenoorth F J.Glutathione-S-transferase and hydrolytic activity in a tetrachlorvinphos resistant strain of house fly and their influence on resistance[J].Pesticide Biochemistry and Physiology,1979,11(1-3),176-178.
[16]唐振华,韩罗珍,张朝远.抗马拉硫磷淡色库蚊不同基因型的自然内禀增长率及其对抗性演化的影响[J].昆虫学报,1990,33(4):385-392.
[17]于金凤,慕立义,王开运.4种棉蚜抗药性种群的生命力及繁殖力[J].植物保护学报,1996,23(1):73-78.
[18]吴孔明,刘芹轩.棉蚜抗杀灭菊酯品系的某些生物学特性[J].昆虫学报,1994,37(2),137-144.
[19]刘凤沂,须志平,薄仙萍,等.昆虫抗药性与适合度[J].昆虫知识,2008,45(3):374-378.
[20]潘文亮,党志红,高占林.棉蚜抗吡虫啉品系和敏感品系主要解毒酶活性比较[J].昆虫学报,2003,46(6):793-796.
[21]吕敏,刘慧霞,吴文军,等.谷胱甘肽-S-转移酶与昆虫抗药性的关系[J].昆虫知识,2003,40(3):204-207.
[22]慕立义,王开运,姜家良,等.棉蚜对不同药剂抗性发生规律及其机理研究[J].农药,1988,27(2):5-7.
