我国大陆甘蔗种质资源利用进展
2012-11-06,,
,,
(广州甘蔗糖业研究所/广东省甘蔗改良与生物炼制重点实验室,广州 510316)
2012-07-14
齐永文(1978-),男,博士,高级农艺师,主要从事甘蔗种质资源与分子育种研究,Email: yongwen2001@163.com。
国家自然科学基金项目(30800700);国家现代农业产业技术体系(CARS-20-1-4);广东省科技计划项目(2011B060400019)。
我国大陆甘蔗种质资源利用进展
齐永文,邓海华,李奇伟
(广州甘蔗糖业研究所/广东省甘蔗改良与生物炼制重点实验室,广州 510316)
我国大陆甘蔗杂交育种始于20世纪50年代。1954/1955杂交季,在海南甘蔗育种场首次实现热带种与本土割手密的杂交。经过50多年的发展,我国大陆甘蔗育成品种的产量、糖分等主要经济性状得到显著改良,种质资源利用的范围显著扩大。综述我国大陆甘蔗育种中主要种质的创新利用进展。
甘蔗;种质资源;中国
甘蔗是中国最为重要的糖料作物,蔗糖占全国食糖总产的90%以上。我国大陆的甘蔗育种始于20世纪50年代,经过近60年的发展,在甘蔗品种改良上取得了显著成效,育出了一系列优良品种,甘蔗品种的产量、糖分都显著提高。种质资源是品种选育的基础,我国一直非常重视甘蔗种质资源的引进和创新研究,不仅从美国、印度、澳大利亚等国家引进了许多优良种质,而且在本土野生种质资源割手密、斑茅的利用研究上也取得显著进展,选育出一批优异种质材料。目前保存在我国甘蔗杂交制种基地——广州甘蔗糖业研究所海南甘蔗育种场的种质资源已有2 100多份,每年该场为全国各育种单位生产2 000个左右的杂交花穗。对我国大陆50年来的甘蔗杂交亲本系谱分析[1]表明,我国甘蔗育种中使用的主要亲本经历了骨干引进亲本为主、自育品种为主以及利用新引进的品种和利用本土野生种质创新为主的3个阶段。随着可用种质数量的不断增加,亲本的遗传基础也在逐渐扩大,尽管目前大部分亲本也还主要是POJ2878、POJ2575的后代,但是各个骨干亲本间的共祖度系数已经显著降低。笔者就我国大陆甘蔗育种上的主要种质利用进行简要回顾与分析,以期为后续的甘蔗种质资源引进、创新和利用提供帮助。
1 中国本土割手密种质资源研究及其在甘蔗育种上的应用
割手密(S.spontaneumL.)是甘蔗近缘野生种之一,具有耐旱、耐贫瘠、抗逆性强等特点,在甘蔗改良中具有重要作用[2]。19世纪初,Jesweit、Barber等通过热带种与割手密的杂交利用,育成了POJ、Co系列高产、高糖种质,为此后的世界甘蔗育种奠定了良好的基础[3,4]。当前,世界范围内种植的甘蔗栽培品种几乎都含有割手密的遗传成分,甘蔗栽培种中约10%的染色体来自于割手密[5]。
中国是割手密的原产地之一,主要分布在北纬18°~33°、东经97°~122°的地区[6]。从20世纪50年代以来,我国科研人员就已开展割手密与热带种、栽培种之间的杂交利用研究。1954/1955杂交季,广州甘蔗糖业研究所在海南三亚首次进行本土割手密与热带种之间的杂交并成功获得杂种后代。此后,通过本土割手密与甘蔗热带种(S.officinarum)、甘蔗杂交种(Saccharumspp.)的连续杂交与回交,选育出了崖城58-43、崖城71-374、崖城84-125等一批优良的具有本土割手密血缘的“崖城”系列材料优良亲本,成为中国大陆甘蔗育种的主要亲本系统之一[7]。截至2011年,由我国本土割手密血缘创新的亲本已育成36个品种。其中源自Badila×崖城割手密组合的崖城58-47和崖城58-43,分别育成后代品种24个和6个,这些品种主要是本土割手密的第3代和第4代杂种[8]。
由于割手密在甘蔗育种中的重要作用,我国科研人员对割手密种质资源的染色体组成、遗传多样性进行了许多研究。Chen 等[9]对中国西南地区割手密的染色体组成进行了分析,结果表明,所分析材料的染色体大部分是2n=80,部分是2n=64,还有极少数是2n=68或2n=116。王水琦等[10]对21份福建“割手密”体细胞染色体数进行的分析表明,割手密的染色体组成包括2n=72、80、84、88、92、96、102等7种类型,也是以2n=80为主。文建成等[11]对来自云南、四川、福建、广东、贵州、江西的106份割手密进行染色体计数观察,染色体数目包括2n=60、64、70、72、78、80、90、92、96、102、104、108共11种类型,可见中国割手密的染色体类型非常丰富。近年来,应用RAPD、SSR、SRAP等分子标记对广东、广西、四川等地的割手密进行了分析[12~14]。这些研究也都表明中国割手密具有丰富的遗传变异,割手密的生长习性受光照、温度等生态因素影响较大,在不同地区形成多种响应生态型。
最近,笔者通过SSR等分子研究表明,由本土割手密创新的亲本育成的品种与从印度、美国、中国台湾等地区引进的种质材料育成的品种具有显著的遗传异质性(未发表),而且这种遗传差异和其所携带的割手密遗传成分存在紧密联系。可见,本土割手密的创新应用,显著拓宽了我国甘蔗品种的遗传基础。
2 斑茅种质资源的开发利用
斑茅(Erianthusarundinaceus(Retz.) Jeswit)是甘蔗近缘属植物之一,具有适应性强、耐旱、抗病虫、宿根性好等优点[15]。文建成等[11]对自云南、 江西、四川、贵州、福建等地的29份斑茅无性系进行染色体数目分析,斑茅的染色体组成全部为2n=60。由于斑茅的高产、高抗逆特性,一直受到甘蔗育种家的重视。广州甘蔗糖业研究所自20世纪50年代起在海南甘蔗育种场开展斑茅的杂交利用研究,于2001年成功获得斑茅蔗第二代(BC1)杂种,此后又获得了BC2及更高世代的回交品系。为了探索斑茅在甘蔗改良上的利用价值,广州甘蔗糖业研究所和澳大利亚甘蔗试验站管理局(BSES)研究人员应用基因组原位杂交技术,对甘蔗与斑茅后代的染色体遗传规律进行了分析[16]。结果表明,在F1(斑茅HN92-77×热带种Badila)代,染色体基本符合n+n的染色体传递方式。在BC1和BC2代基本分别符合2n+n和n+n的染色体传递方式。斑茅与甘蔗杂交、回交过程中,均存在染色体丢失现象,但未观察到染色体发生交换与重组。此后,Deng 等[17]研究认为,BC1中染色体传递比较复杂,既有2n+n模式又有n+n模式。
由于常规杂交过程中,在导入斑茅目的性状的同时,不可避免地导入不良的非目的性状如毛群发达、茎径细等,利用优良品种进行持续回交是减少斑茅所携带的非目的性状的有效方法。符成等[18]对斑茅杂交后代F1、BC1、BC2的主要农艺性状进行了比较分析,如果表明,斑茅与甘蔗杂交的BC2的茎径比F1提高了35%,比BC1提高了10%。BC2的锤度比F1提高了47%,比BC1提高了11%。2006~2007年,笔者对另外一个组合的斑茅蔗BC1、BC2的新植宿根进行试验,也得到相似结果。与BC1群体相比,BC2群体在株高、茎径等主要农艺性状上都得到显著改良,而且有效茎数也没有减少。可见通过连续回交与选择,能够显著改良斑茅带来的不利性状,选育具有目的性状的优良品系。目前,广州甘蔗糖业研究所已经选育出了一批高产、高抗的优良斑茅蔗BC4等高世代回交品系,一些优良的斑茅后代材料已经开始用于甘蔗育种,有望在今后甘蔗品种选育中发挥重要作用。
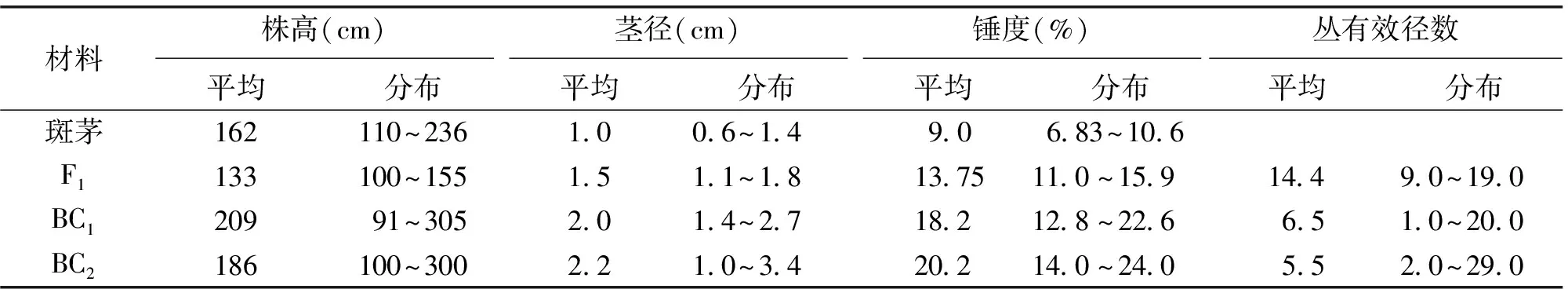
表1 斑茅蔗F1 、BC1 、BC2 主要经济性状比较[18]
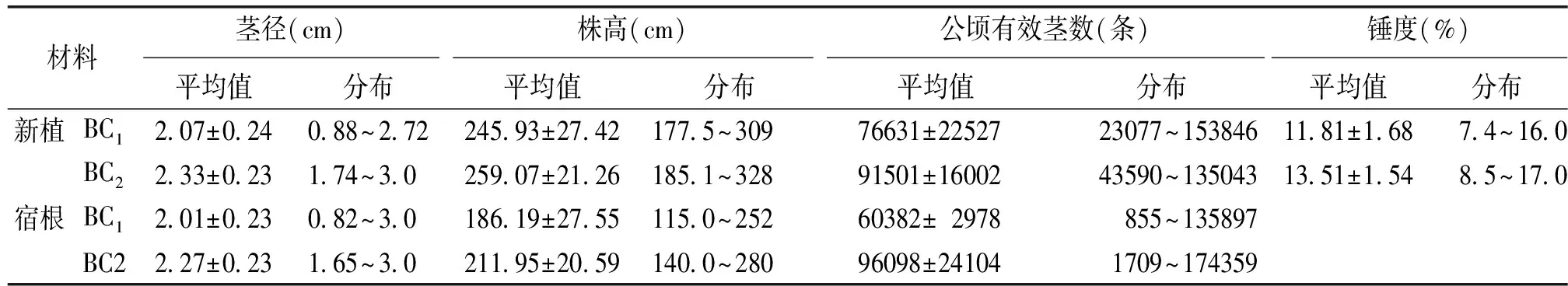
表2 斑茅蔗BC1和BC2主要农艺性状比较
3 国外引进种质的利用
邓海华等[7]对我国大陆自育的175个甘蔗生产品种的亲本统计结果表明,这些品种全部含有国外品种血缘。其中直接利用国外引进品种作亲本(单亲或双亲)育成的品种就有116个(占66.29%)。张琼等[1]对186个有记录的甘蔗品种的亲本进行统计,结果表明,育成这186个品种共使用了93个亲本,其中从国外引进的品种28个,占所有亲本的30.1%。在引进的亲本中来自印度的Co系列品种8个,CP系列品种14个,澳大利亚品种2个,其他国外品种4个。在21个核心亲本(表3)中,有9个国外引进亲本,占核心亲本的42.9%,其中4个来自印度,4个来自美国,1个来自澳大利亚。劳方业等用AFLP标记对36个甘蔗引进品种进行分析,聚类结果表明,36个引进品种为同一大类,而当地亲本材料粤农73-204独立成另一大类,说明中国品种与引进品种之间存在一定的遗传差异。齐永文等[19]应用18对SSR引物对来自美国的12份种质和中国选育的36份种质进行了比较分析。结果表明,一些早期引进的美国材料如CP49-50、CP72-1210、CP33-10等,由于在甘蔗育种中得到广泛利用,已经和中国的品种没有显著遗传差异。一些新引进的品种如CP84-1198、CP94-1100等,与中国甘蔗种质间存在明显的遗传差异,因此,今后应继续加强国外种质尤其是新选育种质的引进和利用。除了美国、印度外,我国还从澳大利亚、巴西、泰国等国家引进了不少种质,但是除极个别的材料在育种中得到应用外,大部分种质还没有得到充分应用,为了提高甘蔗品种的遗传基础,今后对这些材料也应进行重点探索。

表3 我国大陆使用的21个骨干亲本
4 小结与展望
现代甘蔗栽培种是多个种间复合杂交后代,为高度异源多倍体,植株高大,常规杂交育种难度大、周期长。而且由于不同甘蔗品种的遗传背景不一致,开花习性差异很大,导致很多亲本之间无法配制杂交组合。根据广州甘蔗糖业研究所海南甘蔗育种场的记录,从20世纪50年代我国开展甘蔗杂交育种以来,我国实际上常用的甘蔗亲本只有200个左右。能够选育出品种的亲本只有100个左右,而且大部分品种都是来源于少数几个骨干品种。由此可见,优异亲本的选育对于甘蔗育种意义重大。而且,好的品种未必是好的亲本,但一个好的亲本往往能选育出多个优良品种,因此在加强品种选育的同时,更要注重优良亲本的选育和更新,通过扩大种质资源利用范围,为甘蔗育种提供更为丰富的遗传材料。近年来,季节性干旱、霜冻等自然灾害的频繁发生以及甘蔗生产模式的变化对品种特性也提出了更多要求。因此,必须加强种质资源的创新利用,通过源头创新,扩大甘蔗种质资源的利用范围和效率,为培育适应甘蔗产业多元化要求的品种提供遗传基础。
[1] 张 琼, 齐永文, 张垂明, 等.我国大陆甘蔗骨干亲本亲缘关系分析[J].广东农业科学, 2009,(10): 44-48.
[2] Bremer G.Problems in breeding and cytology of sugar cane[J].Euphytica, 1961, 10: 59-78.
[3] D’Hont A, Lu YH, Feldmann P, et al.Cytoplasmic diversity in sugar cane revealed by heterologous probes[J].Sugar Cane, 1993, (1): 12-15.
[4] Grivet L, D' Hont A, Roques D, et al.RFLP mapping in cultivated sugarcane (Saccharumspp.): genome organization in a highly polyploid and aneuploid interspecific hybrid[J].Genetics, 1996, 142: 987-1000.
[5] Sreeinivasan TV, Ahloowalia BS, Heinz DJ. Cytogenetics[A]. In:Heinz DJ.Sugar-cane Improvement Through Breeding[C].Amsterdam: Elsevier,1987.211-253.
[6] Chen SL, Phillips SM.SaccharumLinnaeus, Sp.P1.1:54.1573[A].In:Wu ZY, Raven PH, Hong DY. Flora of China Vol.22 (Poaceae)[C]. Beijing:Science Press; St. Louis:Missouri Botanical Garden Press, 2006.576-581.
[7] 邓海华, 张 琼. 我国大陆近年育成甘蔗品种的亲本分析[J] .广东农业科学,2006,(12):7-10.
[8] 邓海华.我国本土割手密育成品种的亲缘关系分析[J] .广东农业科学, 2012, 39(8): 167-170.
[9] Chen CL, Li LF, Wu SZ.Chromosome number distribution ofSaccharumspontaneumin the southwest region of China[J].International Sugar Journal, 1981, 83: 264-267.
[10] 王水琦, 王子琳, 潘世明, 等.甘蔗种质资源的研究[J].江西农业学报, 2002, 14 (1): 21-26.
[11] 文建成, 蔡 青, 范源洪, 等. 甘蔗属割手密(SaccharumSpontaneum),近缘属斑茅(Sclerostachya)及河八王(Narenga)的染色体数目研究[J].甘蔗糖业, 2001, (3): 12-15.
[12] Chang D, Yang FY, Yan JJ, et al. SRAP analysis of genetic diversity of nine native popula-tions of wild sugarcane,Saccharumspontaneumfrom Sichuan, China[J].Genet Mol Res, 2012, 11(2): 1245-1253.
[13] Zhang GM, Li YR, He WZ, et al. Analysis of the genetic diversity inSaccharumspontaneumL. accessions from Guangxi Province of China with RAPD-PCR[J]. Sugar Tech, 2010, 12(1): 31-35.
[14] 齐永文,樊丽娜,何慧怡,等. 广东割手密资源农艺性状遗传多样性评价[J].甘蔗糖业, 2009,(3):7-10,20.
[15] Besse P, Taylor G, Carroll B, et al. Assessing genetic diversity in a sugarcane germplasm collection using an automated AFLP analysis[J]. Genetica, 1998, 104: 143-153.
[16] Piperidis N, Chen JW, Deng HH, et al. GISH characterization ofErianthusarundinaceuschromosomes in three generations of sugarcane intergeneric hybrids[J]. Genome, 2010, 53(5): 331-336.
[17] Deng ZH, Zhang MQ, Lin W, et al. Analysis of disequilibrium hybridization in hybrid and backcross progenies ofSaccharumofficinarum×Erianthusarundinaceus[J].Agricultural Sciences in China, 2010, 9(9): 1271-1277.
[18] 符 成, 邓海华, 杨业后, 等.斑茅及其杂种后代主要经济性状研究[J].甘蔗糖业, 2004, (1):1-5.
[19] 齐永文, 劳方业, 张垂明, 等.中美重要甘蔗种质SSR遗传多样性比较[J].热带作物学报, 2011, 32(1): 99-104.
AdvanceinUtilizationofSugarcaneGermplasminChinaMainaland

(Guangzhou Research Institute for Sugarcane Industry/Guangdong Key Lab of Sugarcane Improvement and Biorefine, Guangzhou, Guangdong 510316, China)
Crossbreeding of sugarcane in China mainland began in the 1950s. In 1954-1955, interspecific cross between S.officinarumand native S.spontaneumfirstly succeed in Hainan Sugarcane Breeding Station. Through development for over 50 years, major economic traits of bred varieties, such as yield, sugar content, were significantly improved, and the range of utilization of Germplasm was significantly expanded. This paper reviewed the advance in utilization of main sugarcane germplasms in China mainland sugarcane breeding program, to provide help for related research.
Sugarcane; Germplasm; China
S566.102
A
1001-5280(2012)05-0443-04
10.3969/j.issn.1001-5280.2012.05.08
