河北沿岸微微型浮游植物的分布特征
2012-10-23李洪波刘述锡林凤翱冯志权
李洪波, 刘述锡, 林凤翱, 冯志权
(国家海洋环境监测中心, 辽宁 大连 116023)
河北沿岸微微型浮游植物的分布特征
李洪波, 刘述锡, 林凤翱, 冯志权
(国家海洋环境监测中心, 辽宁 大连 116023)
于2006年7月~ 2007年10月间, 分4个季度调查了河北省沿岸微微型浮游植物的丰度和生物量及对浮游植物总生物量的贡献。结果显示: 河北近岸海域聚球藻蓝细菌丰度为 4.46×103个/mL(0.79×103~ 16.19×103个/ mL), 生物量(以碳计, 下同)为 1.31 mg /m3(0.84~17.47 mg /m3), 季节分布特征为秋季>冬季>夏季>春季。微微型光合真核生物丰度为 4.43×102个/mL (0.84×102~ 17.47×102个/mL), 生物量为1.11mg /m3(0.21~ 4.37 mg/m3), 季节变化变现为秋季>冬季>春季>夏季。微微型浮游植物对浮游植物总生物量的贡献年平均为5.32%(1.84%~ 8.91%), 春季最高, 秋季最低。温度在较冷季节(冬春季)里是影响聚球藻蓝细菌生长和分布的控制因素。总之, 在近岸环境里, 微微型浮游植物并不占优势。
河北沿岸; 微微型浮游植物; 分布; 荧光显微镜
海洋微微型浮游植物(picophytoplankton, 0.2~2µm), 主要包括聚球藻蓝细菌(Synechococcus spp.,简称SYN)、原绿球藻(Prochlorococcus, 简称PRO)和微微型真核藻类(Pico-eukaryote, 简称 PEUK), 它们存在于世界海洋的大多数海区中, 是浮游植物的重要组成部分[1]。而在近岸海域, 由于多为富营养条件,原绿球藻则较少分布[2]。对海洋微微型浮游植物的生态作用及研究方法国内已有报道[3-4], 微微型浮游植物广泛分布于世界各个纬度, 对世界海洋初级生产力和浮游植物生物量的贡献分别达40%和80%[5-6]。
国际海洋微微型浮游植物的研究始于 20世纪70年代末[7], 中国从 90年代初期开始, 对微微型浮游植物在中国海域的生态分布得到广泛的关注[8-21]。但在近岸海域有关微微型浮游植物则报道较少。本项目在国家908的支持下, 于2006年7月~ 2007年10月间, 分4个季度来调查微微型浮游植物在河北近岸环境中的分布, 由此来说明其在近岸环境中的作用。
1 采样和方法
1.1 采样站位和采样时间
调查时间为2006年7月~2007年10月, 每季度调查一次, 时间分别为 2006年7月(夏季)、2006年11月(冬季)、2007年 4月(春季)、2007年 10月(秋季)。采样站位如图1所示。水深超过10 m, 按照表、中、底采样; 水深小于10 m, 按照表、底采样。站位共计27个, 其中水深大于10 m的站位有15个。
1.2 微微型浮游植物的计数和数据处理
微微型浮游植物的计数方法按照《海洋调查规范》[22]进行。取5~20 mL水样, 经孔径为0. 2 μm的黑色核孔滤膜过滤后, 制片。在荧光显微镜下用绿光和蓝光激发, 使用40倍物镜观察, 随机取至少20个视野, 分别计数具有光亮的橘黄色荧光的含藻红蛋白的 Syn细胞和呈砖红色荧光的含叶绿素的 PEUK细胞。
1.3 微微型浮游植物丰度和生物量的计算
丰度计算方法参照《生物生态调查技术规程》。聚球藻蓝细菌和真核微微型光合生物生物量的换算,分别按照每个细胞含有 294 fgC[23]和 2500 fgC[24]来估算。
叶绿素 a与浮游植物生物量之间的转换关系按照1 单位叶绿素a等于45单位碳含量[25]来换算将叶绿素a 浓度转换为浮游植物生物量(mg /m3)。

图1 调查海区及采样站位Fig. 1 Samping stations in the coastal areas of Hebei
用Sufer8. 0 软件分析各季节水层微微型浮游植物的平面分布趋势。
2 结果和讨论
2.1 聚球藻蓝细菌的时空分布2.1.1 聚球蓝细菌的水平及季节分布
2006年7月(夏季), 聚球藻蓝细菌的平均丰度为4.12×103个/mL(1.56×103~9.14×103个/mL), 最小值出现在HBBZ01底层, 最大值出现在HBBZ02中层;2006年 11月(冬季), 聚球藻蓝细菌的平均丰度为5.45×103个/mL(1.15×103~14.73×103个/mL), 最小值出现在 HBBZ08底层, 最大值出现在 BD06表层;2007年 4月(春季), 聚球藻蓝细菌的平均丰度为2.57×103个/mL(0.79×103~6.47×103个/mL), 最小值出现在HBBZ09中层, 最大值出现在HBBZ04中层;2007年 10月(秋季), 聚球藻蓝细菌的平均丰度为5.68×103个/mL(1.31×103~16.19×103个/mL), 最小值出现在 ZD-BDH064底层, 最大值出现在ZD-BDH076底层(表 1)。聚球藻蓝细菌的生物量见表 1。

表1 河北沿岸微微型浮游植物的丰度和生物量Tab. 1 Abundance and biomass of picophytoplankton along Hebei coastal area
从图 2表层蓝细菌水平分布来看, 在夏季和秋季滦河口附近出现丰度较小值外, 其他两季节在滦河口出现最大值。这可能与夏秋季降水有关, 河流携带泥沙, 影响表层蓝细菌的分布。除夏季外, 蓝细菌在河北沿岸分布呈北高南低趋势。沿离岸方向, 其丰度逐渐降低。
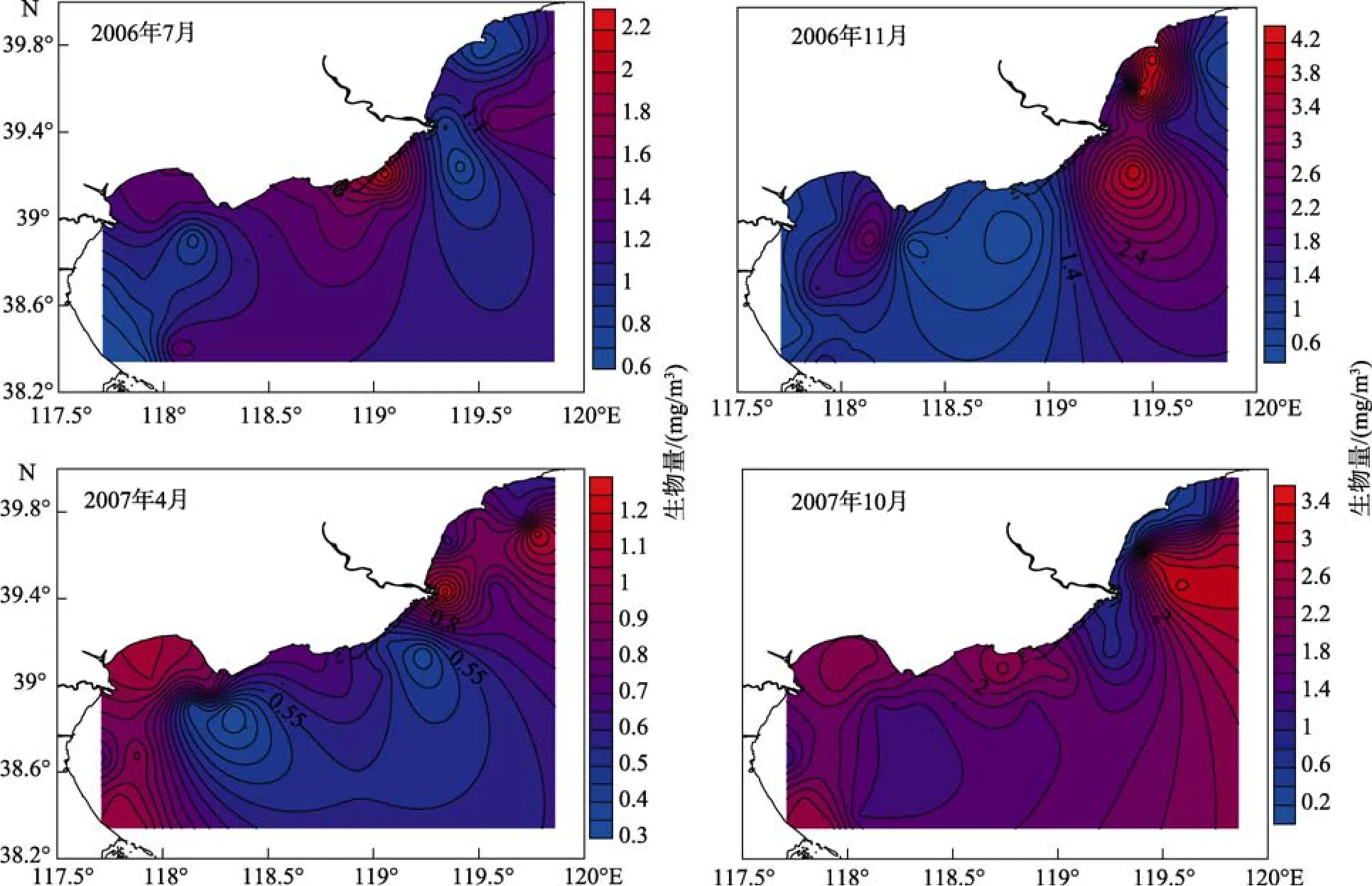
图2 河北沿岸区域表层聚球藻蓝细菌生物量分布概况Fig. 2 Distribution of Synchococcus biomass in Hebei coastal surface waters
全年蓝细菌 丰度变化 从 0.79×103~16.19×103个/mL, 平均值为 4.46×103个/mL; 全年生物量0.84~17.47 mg/m3, 平均值为1.31 mg/m3。季节分布特征为秋季>冬季>夏季>春季。
2.1.2 聚球蓝细菌的垂直分布
从表 2得知, 整个海区表层聚球藻蓝细菌年平均丰度为 4.52×103个/mL(n=109), 中层平均丰度为4.73×103个/mL(n=59), 底层平均丰度为 4.23×103个/mL(n=102), 河北沿岸海域年蓝细菌平均值在垂直方向上表现为中层>表层>底层。但秋冬季聚球藻蓝细菌在表层最高, 最低值分别出现在中层和底层。秋季和冬季可能受季风影响, 同时受光照影响, 聚球藻在表层较大。
2.2 微微型真核藻类的时空分布
2.2.1 微微型真核藻类的水平及季节分布
2006年7月(夏季), 微微型真核藻类的平均丰度为 1.64×102个/mL(0.84×102~9.70×102个/mL), 最小值出现在HBBZ07表层, 最大值出现在HBBZ02中层; 2006年11月(冬季), 平均丰度为6.11×102个/mL(1.37×102~17.47×102个/mL), 最小值出现在HBNZ07表层, 最大值出现在ZD-TJ077中层; 2007年4月(春季), 平均丰度为 3.43×102个/mL(1.26×102~8.89×102个/mL), 最小值出现在 HBBZ04底层, 最大值出现在HBBZ06中层; 2007年10月(秋季), 平均丰度为6.54×102个/mL(1.82×102~13.78×102个/mL), 最小值出现在ZD-BDH064底层, 最大值出现在HBNZ05表层(表2)。微微型真核藻类的生物量见表1。

表2 4个季度垂直方向上的微微型浮游植物分布概况Tab. 2 The distribution of pico-phytoplankton along vertical direction
从图3表层微微型光合真核藻类水平分布来看,河北沿岸秋冬季分布呈北高南低趋势; 春夏季节分布正好相反, 春季为中间高, 两侧低; 而夏季为两侧高, 中间低。沿离岸方向, 其丰度有下降趋势。

图3 河北沿岸表层微微型真核浮游植物生物量分布Fig. 3 Distribution of pico-eukaryote biomass in Hebei coastal surface water
全年微微型光合真核藻类丰度变化从0.84×102~17.47×102个/mL, 全年平均值为 4.43×102个/mL; 全年生物量为0.21~4.37 mg/m3, 平均值为1.11 mg/m3。季节变化变现为秋季>冬季>春季>夏季。
与聚球藻蓝细菌相比,可得出蓝细菌丰度约为微微型光合真核藻类丰度的 10倍(蓝细菌丰度为4.46×103个/mL, 微微型光合真核藻类丰度 4.43×102个/mL)。
2.2.2 微微型真核藻类的垂直分布
从表 2知道, 整个海区微微型真核藻类全年的表层平均丰度为 4.58×102个/mL(n=109), 中层平均丰度为 4.80×102个/mL(n=59), 底层平均丰度为4.07×102个/mL(n=102), 故河北沿岸微微型真核藻类垂直变化为中层>表层>底层, 但各层相差不大。而秋季微微型真核藻类的垂直分布表现为表层>底层>中层, 秋季微微型真核藻类在表层值较大, 可能是受季风、光照和营养盐协同影响的结果。
从表 3列出的中国沿海微微型浮游植物丰度的分布来看, 聚球藻丰度在 103~105个/mL, 微微型真核藻类丰度在 102~104个/mL。相比其他海区, 聚球藻和微微型真核藻类在河北沿岸的丰度较低。
2.3 微微型浮游植物对总生物量的贡献
河北近岸的叶绿素a浓度和生物量见表4。从中可以看出叶绿素 a季节分布表现为夏秋季高, 春冬季低。春季较低的原因可能与调查时间偏早有关, 水温偏低(春季表层水温平均小于 9℃), 制约了浮游植物的生长。
表 5 列出了河北沿岸微微型浮游植物生物量对浮游植物总生物量的贡献。从中可以得到, 暖季节(夏秋季), CB:PB 和 EB:PB 都较低, 其中秋季最低;而冷季节(冬春季), CB:PB和EB:PB都较高, 其中春季最高。PPB:PB在4个季度表现为春季>冬季>夏季≈秋季, 春季平均比值大概是夏季的5倍。微微型浮游植物对总生物量的贡献年平均为 5.32%(1.84%~8.91%), 此结果远远小于作者曾在黄海冷水团的调查结果[21], 在黄海冷水团中由微微型聚球藻蓝细菌对浮游植物总生物量的贡献平均达42.5%。本调查结果与肖天[12]在渤海的结果较为一致, 聚球藻蓝细菌生物量在总浮游植物生物量(CB/PB) 中占的比例秋季平均为6.4%, 春季平均为0.3%; 在东海调查结果为 CB/PB冬季平均是 10%, 夏季平均是3%[13]。

表3 中国近海微微型浮游植物的分布Tab. 3 The abundance of picophytoplankton in coastal area of China

表4 叶绿素a浓度及浮游植物总生物量Tab. 4 Concentration of Chl-a and phytoplankton biomass in four seasons

表5 微微型浮游植物与浮游植物总生物量的比值Tab. 5 Ratio of pico-phytoplankton biomass to phytoplankton biomass
河北沿岸区域的微微型浮游植物的生物量占浮游植物总生物量的比重较低, 说明在近岸海域, 微微型并不是浮游植物的主要类群。
2.4 微微型浮游植物与温度的关系
针对微微型浮游植物中的原核生物——聚球藻蓝细菌的生物量在四个季度与表层水温的关系进行分析。把全年分为冷季节(春季和冬季平均表层海洋温度(surface sea temperature) SST分别为8.9℃和7.4℃)和暖季节(夏季和秋季平均 SST分别为 23.6℃和18.2℃)。SST 在夏秋季平均为 21℃ (16.2 ~25.2℃)。
从图 4分析得到, 在温度超过 15℃, 聚球藻蓝细菌生物量随温度升高而降低。Li[26]对许多研究聚球蓝细菌和异养细菌的文献数据进行回归, 认为在较暖的水域里(>14℃), 随温度升高, 蓝细菌生物量有降低趋势, 说明其他比如底质供应(substrate supply)、营养盐等影响着蓝细菌的生长以及分布。在平均温度低于 14℃的情况下, 聚球蓝细菌的平均丰度直接与温度相关, 且相关性较高。在较冷季节里, 温度是影响细菌生长和分布的控制因素。

图4 夏、秋季聚球藻蓝细菌生物量与表层水温的关系Fig. 4 The relationship between cyanobacterial biomass and surface sea temperature (SST) in summer and autumn
3 结论
微微型浮游植物有明显的季节改变。在河北近岸海域, 微微型浮游植物的丰度秋季最高, 春季最低。
微微型浮游植物对浮游植物总生物量的贡献年平均为5.32%(1.84%~8.91%), 春季最高, 秋季最低。
在较冷季节里, 温度是影响微微型聚球藻蓝细菌生长的重要因素。
[1] 宁修仁. 海洋微型和超微型浮游生物[M]//苏纪兰,秦蕴珊. 当代海洋科学学科前沿. 北京: 学苑出版社,2000: 272-277.
[2] 焦念志, 杨燕辉. 中国海原绿球藻研究[J]. 科学通报,2002, 47(7): 485-491.
[3] 肖天, 李洪波, 赵三军, 等. 海洋浮游细菌在碳循环方面的作用[J]. 海洋科学, 2004, 28(9): 46-49.
[4] 孙晓庆, 董树刚. 微微型浮游植物的生态学研究进展[J].海洋科学, 2008, 32(5): 67-72.
[5] Li W K W, Subbarao D V, Harrison W G, et al. Autotrophic picoplankton in the tropical ocean [J]. Science,1983, 219: 292-295.
[6] Murphy L S, Haugen E M. The distribution and abundance of phototrophic ultraplankton in the North Atlantic [J]. Limnol Oceanogr, 1985, 30: 47-58.
[7] Waterbury J B, Watson S W, Guillard R, et al. Widespread occurrence of a unicellular, marine planktonic cyanobacterium[J]. Nature, 1979, 277: 293-294.
[8] 陈怀清, 钱树本. 青岛近海及其邻近海域微型、超微型浮游藻类的研究[J]. 海洋学报, 1992, 14(3):105-113.
[9] 屈佩, 张学雷, 王宗灵,等. 南黄海夏季微微型浮游植物丰度的分布[J]. 海洋学报, 2010, 32(4): 155-167.
[10] 宁修仁. 长江口及其毗邻东海水域蓝细菌的分布和细胞特性及其环境调节[J]. 海洋学报,1991, 15(4):552-559.
[11] 宁修仁, 蔡昱明, 李国为, 等. 南海北部微微型光合浮游生物的丰度及环境调控[J]. 海洋学报, 2003,25(3): 83-97.
[12] 肖天, 王荣. 春季与秋季渤海蓝细菌(聚球蓝细菌属)的分布特点[J]. 生态学报, 2002, 220(12): 2071-2078.
[13] 肖天, 岳海东, 张武昌, 等. 东海聚球蓝细菌(Synechococcus)的分布特点及在微食物环中的作用[J].海洋与湖沼, 2003, 34(1): 33-43.
[14] 黄邦钦, 洪华生, 林学举. 台湾海峡微微型浮游植物的生态研究. I 时空分布及其调控机制[J]. 海洋学报,2003, 25(4): 72-82.
[15] 汪岷, 梁彦韬, 白晓歌, 等. 北黄海夏季微微型浮游植物的分布[J]. 植物生态学报, 2008, 32(5):1184-1193.
[16] 汪岷, 梁彦韬, 白晓歌, 等. 青岛近海及其邻近海域夏季微微型浮游植物丰度的分析[J]. 中国海洋大学学报, 2008, 38(3): 413-418.
[17] 汪岷, 梁彦韬, 白晓歌, 等. 青岛近海及其邻近海域冬季微微型浮游植物的分布[J]. 应用生态学报,2008 , 19 (11): 2428-2434.
[18] 肖天, 焦念志, 王荣. 胶州湾蓝细菌、异养细菌的数量分布特点[A]. 胶州湾生态学研究[C]. 北京: 科学出版社, 1995, 118-124.
[19] 孙晟, 肖天, 岳海东. 秋季与春季东、黄海蓝细菌(Synechococcus spp. )生态分布特点[J]. 海洋与湖沼,2003, 34(2): 161-168.
[20] 赵三军, 肖天, 李洪波, 等. 胶州湾聚球菌(Synechococcus spp. ) 蓝细菌的分布及其对生产力的贡献[J] . 海洋与湖沼, 2005, 36(6): 534-540.
[21] Li H B, Xiao T, Ding T, Lv R H. Effect of the Yellow Sea Cold Water Mass (YSCWM) on distribution of Bacterioplankton[J]. Acta Ecologica Sinica, 2006,26(4): 1012-1020.
[22] 中华人民共和国国家质量监督检疫总局. GB/T 12763.9-2007 海洋调查规范[S]. 北京: 中国标准出版社, 2007.
[23] Cuhel R, Waterbury J B. Biochemical composition and sho rt term nutrient incor- poration patterns in unicellular marine cyanobacterium, Synechococcus(WH7803)[J]. Limnol Oceanogr, 1984, 29: 370-374.
[24] Liu H B, Suzuki k, Minami C, et al. Picoplankton community structure in the subarctic Pacific Ocean and the Bering Sea during summer 1999[J]. Mar Ecol Prog Ser, 2002, 237: 1-14.
[25] Krempin D W, Sullivan C W. The seasonal abundance,vertical distribution, and relative microbial biomass of chrococcoid cyanobacteria at a station in southern California coastal waters[J]. Can J Microbiol, 1981, 27:1341-1344.
[26] Li W K W. Annual average abundance of heterotrophic bacteria and Synechococcus in surface ocean waters[J].Limnol Oceanogr, 1998, 43(7): 1746-1753.
Dsistribution characteristics of picophyplankton in coastal area of Hebei
LI Hong-bo, LIU Shu-xi, LIN Feng-ao, FENG Zhi-quan
(National Marine Environmental Monitoring Center, Dalian 116023, China)
Jun.,14,2011
coastal area of Hebei; picophytoplankton; distribution; epifluorescence microscopy (EFM)
From July 2006 to October 2007, the picophytoplankton abundance, biomass and the contribution of picoplankton to phytoplankton biomass were conducted seasonly in coastal area of Hebei. The results showed that the average abundance of cyanobacteria Synechococcus spp. was 4.46×103cells/mL (0.79×103to 16.19 ×103cells/mL);the average biomass was 1.31 mg/m3(from 0.84 mg/m3to 17.47 mg/m3). The seasonal variable of Synechococcus spp.was autumn>winter>summer>spring. The range of eukaryotic picophytoplankton abundance was from 0.84×102cells/mL to 17.47×102cells/mL (on average 4.43×102cells/mL), and the biomass was from 0.21 mg /m3to 4.37 mg/m3(average 1.11 mg/m3). The seasonal variable of pico-eukaryotic phytoplankton was autumn>winter >spring>summer. The contribution of picophytplankton to total phytoplankton biomass (PPB:PB) was from 1.84 % to 8.91 % (average vaule was 5.32 %), which was lower than offshore and oligotrophic water. In colder season, temperature was an important factor that affecting the growth and distribution of picophytoplankton. In a word, picophytoplankton was not the dominant phytoplankton in coastal area of Hebei.
P722.5
A
1000-3096(2012)07-0049-07
2011-06-14,
2011-12-15
河北省 908专项(908-HB-02); 国家自然科学基金项目(40906082); 国家海洋局沿岸海域生态环境重点实验室开放基金项目(200903)
李洪波(1976-), 男, 河南漯河人, 博士, 助理研究员, 主要从事海洋微型生物生态与环境研究, 电话: 0411-84782711, E-mail:hbli@nmemc.gov.cn
(本文编辑:梁德海)
