3种配方处理吉贝插穗生根过氧化物酶(POD)活性变化的研究
2012-09-20伍建榕马焕成
高 柱,伍建榕,严 毅,郑 元,马焕成*
(1.西南林业大学国家林业局西南地区生物多样性保育重点实验室,昆明 650224;2.江西省科学院生物资源研究所,南昌 330029)
山地棉是指在山地上种植具有较好适应性,以生产纺织纤维原料为主,具有较高经济效益的新型天然纤维原料类植物的总称,包括木棉科(Bombacaceae)部分植物、牛角瓜(Calotropis gigantean L.)、长生棉(Gossypium peruriaxum)等。山地棉植物的纤维具有中空度高、纺纱强度高、抗菌、光洁等优良特性。棉花是重要的战略储备物资,我国近20年来种植面积下降0.1%,山地棉种植开发能够有效缓解棉粮争地矛盾。木棉纤维中空度高达86%以上,是人工纤维的2倍,具有光洁、抗菌、轻柔、保暖等优点,吉贝[Ceiba pentandra(Linn.)Gaertn.]是山地棉木棉科植物中纤维开发价值较高的树种之一[1-2]。
扦插育苗具有繁殖系数高、可工厂化育苗等优点,是众多树种无性繁殖育苗的首选[3]。林秀香采用沙壤土∶谷壳灰=4∶1基质扦插1年生吉贝半木质化枝条,以初夏梅雨季节成活率最高[4];本实验前期研究发现,以纯净河沙扦插吉贝,生根率依次为配方1#>配方3#>配方2#(1~3#配方主要成分依次为生根粉、吲哚乙酸和萘乙酸),且均达到70%以上,但有关吉贝扦插过程中生理生化指标变化规律的研究尚未见报道。过氧化物酶(Peroxidase,POD)普遍存在于植物体内,参与多种生理生化过程,与植物不定根的发生和生长有密切关系,在植物细胞生长、分化调节及发育进程中起着重要作用[5-6],Garspar等认为POD活性可作为插穗生根的标志之一[7]。本试验比较了不同专用配方处理对吉贝插穗生根过程中POD活性变化响应规律,为揭示吉贝扦插生根机理提供科学依据。
1 试验地概况
试验在西南林业大学林学院教学试验温室大棚进行。该地位于云南省昆明市盘龙区,地处102°45′16.7″E,25°03′53.9″N,海拔1 959 m。年均温度15℃,积温4 522.6℃,最热月平均气温19℃,最冷月平均气温8℃;昼夜温差大,春、冬两季温差可达12~20℃,紫外线强,光照充足,年均日照时数2 054 h。干、湿季节分明,5~10月为雨季,降雨量占全年85%。试验棚内最低温18℃,最高温48℃,平均气温28℃,最高相对湿度为85%,最低相对湿度为45%,平均相对湿度为80%。
2 材料与方法
2.1 插穗选取及制备
试验于2010年3月7日进行,选取采穗圃地2年生母树的当年生枝条制穗。将剪取的枝条去除叶片,上切口离芽1.0 cm平切、下切口离节0.5 cm处斜切,插穗长20 cm。用石蜡涂抹上切口,剪好的插穗每20根为一捆,用800倍多菌灵溶液浸泡20 min,晾干后备用。
2.2 插床建立及实验设计
采用配有全光照自动间歇喷雾装置的温室育苗池育苗,插床长6 m,宽1.0 m,深40 cm,双面砌墙水泥面,床底铺1层10 cm厚的大鹅卵石,上层铺30 cm纯净河沙。用50%多菌灵可湿性粉剂1 000倍液消毒后,用薄膜覆盖,1周后扦插。分别采用专用配方1#、2#、3#处理插穗1 h。完全随机区组设计,每个处理80根插穗,3次重复,清水处理为对照。扦插深度为穗长2/3,密度50根·m-2。保持河沙湿润,每周消毒1次(800倍多菌灵和甲基托布津交替使用)。及时清除枯穗,新叶完全展开后每隔15 d喷1次2‰尿素。扦插后第0、7、14、21、28、35天取样,每区随机抽取5根插穗,洗净后,观察插穗形态变化,并迅速剥取插穗基部2 cm内韧皮部,超低温冰箱保存,备用。
2.3 POD活性测定方法
称取样品0.1 g,加少许石英砂和液氮研磨,研细后加入2 mL磷酸缓冲液研磨成匀浆,转入离心管,再用2 mL磷酸缓冲液分2次冲洗研钵,并入离心管。10 000 r·min-1冷冻离心15 min,上清液即为酶粗提取液。POD活性测定采用愈创木酚染色法,470 nm波长测定OD值,每分钟读数1次,活性以ΔOD·g-1·min-1表示[8]。
2.4 数据分析
试验数据采用SPSS 16.0进行双变量相关性和单因素方差分析,制图采用Excel 2007软件进行。
3 结果与分析
3.1 插穗生根过程外部形态观察
3种配方处理吉贝插穗生根外部形态观测结果表明,扦插第0~7天,下切口皮层、韧皮部、木质部开始松动,并在木质部外围形成一圈间隙,愈伤组织就在插穗切口形成的间隙处沿韧皮部形成,为乳白色半透明状(见图1-A),有些愈伤组织则在髓芯产生(见图1-B);第7~14天,愈伤组织继续增多,形成白色突起(见图1-B),此时认为已有根原基分化[3],随着扦插时间的延长,不定根形成;第14天以后,大量乳白色不定根突破愈伤组织,不定根数量逐渐稳定(见图1-C);不定根继续伸长,乳白色不定根逐渐木质化,形成灰褐色成熟根系(见图1-D)。因此,吉贝插穗生根可分为3个阶段:0~7 d为愈伤组织形成期,7~14 d为不定根形成期,14 d以后为不定根伸长期。
3.2 三种配方处理插穗POD活性变化规律
由图2-A、B、C可知,3种配方处理吉贝插穗生根过程中,0~21 d时POD活性均呈现“升高-下降-升高”趋势,与CK趋势较相似,其中峰值出现在第14天,大小依次为专用配方3#(55.39 ΔOD·g-1·min-1)、配方 1#(52.29 ΔOD·g-1·min-1)、配方 2#(44.992 ΔOD·g-1·min-1)、 CK(23.96 ΔOD·g-1·min-1),此时经历愈伤诱导期和不定根形成期,至不定根生长期,POD活性开始下降。第21~28天,配方1#的POD活性先降后升;配方2#持续降低,而配方3#则保持上升趋势。配方处理吉贝插穗后,能够显著提高生根过程中7~21 d时插穗的POD活性,并促进生根。

图1 吉贝插穗生根过程中的形态变化Fig.1 Morphological changes of Ceiba pentandra cuttings during rooting process
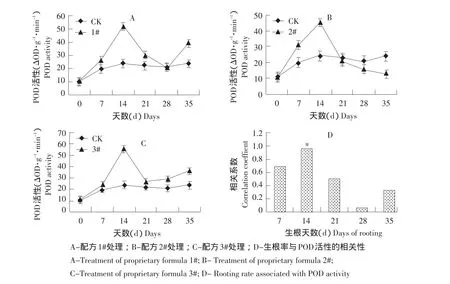
图2 吉贝插穗生根过程中POD活性变化及相关性分析Fig.2 Changes of POD activity and correlation analysis in Ceiba pentandra cuttings during rooting process
将3种配方处理吉贝插穗生根率与生根过程中POD活性进行相关性分析(见图2-D)可知,生根率与不定根形成期(第14天或峰值)的POD活性呈显著正相关(P=0.038),相关系数r=0.962,而与其他时期POD活性差异不显著。说明POD活性越大,对不定根的形成越有利,并最终提高生根率。
3.3 三种配方处理插穗生根期间POD活性变化比较
将图2-A、B、C数据进行方差分析表明,三种配方处理后在不同天数间POD活性变化差异达显著水平(见表1),说明POD活性与吉贝生根密切相关。3种配方处理都极大影响POD活性,但同一天数时不同配方间差异不显著,说明在吉贝插穗生根过程中,3种配方影响POD活性进而促进生根有近似的生理作用。
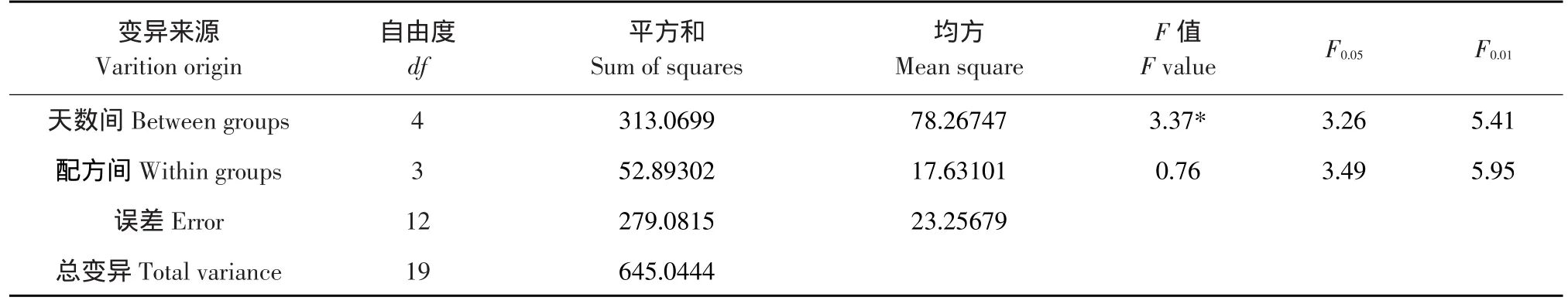
表1 3种配方对POD活性作用的方差分析Table1 Variance analysis of the effect of three kinds of proprietary formula on POD activity
4 讨论与结论
4.1 配方处理对吉贝扦插生根阶段的划分
按照不定根在插穗上着生部位不同,可将插穗生根分为皮部生根、愈伤组织生根和混合生根3种类型。吉贝插穗生根先形成愈伤后有不定根,因此,属于愈伤组织生根型。李明等认为易生根的桉树生根过程可分为愈伤组织诱导期、不定根形成期、新根伸长期[9];难生根的四倍体刺槐硬枝扦插生根分为愈伤诱导期、根原基诱导期和不定根表达与生长期[3];草本芍药不定根形成包括诱导期、表达期和伸长期[10-11]。本试验3种配方处理吉贝插穗扦插生根可分为愈伤组织形成期(0~7 d)、不定根形成期(7~14 d)、不定根伸长期(14 d以后)3个阶段,与前人的研究结果类似。
4.2 吉贝生根与POD酶活性变化的关系
POD是一类含铁卟啉辅基的氧化酶,普遍存在于植物体内,其同工酶在植物体内具有发育阶段特异性和组织特异性,是一种重要的生长调节酶和生根关联酶[12-13],在植物生长发育过程中有重要作用,对植物器官形态建成有特殊作用[14]。在生根过程中,POD多种同工酶中除具有清除活性氧外,也能行使吲哚乙酸氧化酶(IAAO)活性的作用氧化IAA,从而降低插穗体内IAA浓度,促进根原基的诱导[15-16]。宋金耀等研究认为毛白杨嵌合体扦插生根POD活性有2个峰[17],扈红军等发现榛子插条生根过程POD活性仅1个峰[18],而难生根树种四倍体刺槐硬枝扦插生根过程中,POD活性持续呈上升趋势[3],可见POD活性变化的复杂性及关联度的差异性。本研究发现,POD活性在吉贝扦插过程中表现出相似的变化响应,在生根前两个阶段,3种配方处理插穗POD活性均呈现“升高-下降”趋势,活性增加幅度依次为配方3#、配方1#、配方2#,愈伤组织及不定根形成期POD活性上升,能够抑制伤呼吸和降低自由基的损伤,保护细胞膜系统,并抑制插穗变褐腐烂[16],POD活性增加是插穗逆境反应酶之一。通过相关性分析还发现,POD活性最高时与吉贝生根率呈显著正相关,POD活性大小与吉贝生根关系紧密。进一步方差分析显示,在扦插进程中POD活性差异显著,而不同处理间差异不显著,说明POD活性与吉贝插穗生根密切相关,且配方处理间存在相似的作用,该结果与黄卓烈等研究结论相似[19]。此外,过氧化物酶(POD)为吉贝生根重要关联酶,且在愈伤诱导期和不定根形成期需维持较高活性。
[1]李文华,熊定国.木棉产业导引——首届中国木棉产业发展研讨会文集[C].北京:中国纺织出版社,2009:10.
[2]高柱,王小玲,汪洋,等.木棉栽培技术研究进展[J].江西科学,2009,27(5):761-766.
[3]王小玲,赵忠,权金娥,等.外源激素对四倍体刺槐硬枝扦插生根及其关联酶活性的影响[J].西北植物学报,2011,31(1):116-122.
[4]林秀香.青皮木棉引种试种初报[J].热带林业科学,2007,27(1):12-14.
[5]王小玲,高柱,余发新,等.观赏羽扇豆离题培养生根关联酶活性及可溶性蛋白含量变化规律研究[J].云南农业大学学报,2010,25(6):835-389.
[6]曹帮华,扈红军,张大鹏,等.桑树硬枝扦插生根能力及其生根关联酶活性的研究[J].蚕业科学,2008,34(1):96-100.
[7]Gaspar T,Kevers C,Hausman J F,et al.Practical uses of peroxidase activity as a predictive marker of rooting performance of micropropagated shoots[J].Agronomie,1992(12):757-765.
[8]张治安,陈展宇.植物生理学实验技术[M].吉林:吉林大学出版社,2008:7.
[9]李明,黄卓烈,谭绍满,等.难易生根桉树的过氧化物酶活性及同工酶多型性比较研究[J].华南农业大学学报,2000,21(3):56-59.
[10]Hausman J F,Evers D,Kevers C,et al.Internal controls of root induction in poplar shoots raised in vitro[J].Angew Bot,1997,71:104-107.
[11]付喜玲,郭先峰,康晓飞,等.IBA对芍药扦插生根的影响及生根过程中相关酶活性的变化[J].园艺学报,2009,36(6):849-854.
[12]Moneousin C H,Gaspar T.Peroxidase as a marker for rooting improvement in Cynara seolymus L.cultivated in vitro[J].Bioehem Physiol Pflanzen,1983,178:263-271.
[13]Nordstrom A C,Jacobs F A,Eliasson L.Effect of exogenous indole-3-acetic acid and indole-3-butyric acid on internal levels of the respective auxins and their conjugation with aspartic acid during adventitious root formation in pea cuttings[J].Plant Physiol,1991,96:856-861.
[14]潘健.三种柃木属植物扦插生根机理研究[D].南京:南京林业大学,2007.
[15]原牡丹,侯智霞,翟明普,等.IAA分界代谢相关酶(IAAO、POD)的研究进展[J].农业生物技术科学,2008,24(8):88-92.
[16]高柱,王小玲,余发新,等.金边瑞香插穗形态、POD与可溶性蛋白含量变化研究[J].东北农业大学学报,2009,41(4):33-37.
[17]宋金耀,何文林,李松波,等.毛白杨嵌合体扦插生根相关梨花特性分析[J].林业科学,2001,37(5):64-67.
[18]扈红军,曹帮华,尹伟伦,等.榛子嫩枝扦插生根相关酶活性变化及繁殖技术[J].林业科学,2008,44(6):59-65.
[19]黄卓烈,李明,詹福建,等.不同生长素处理对桉树无性系插条氧化酶活性影响的比较研究[J].林业科学,2002,38(4):46-52.
