围栏封育对青藏高原东缘高寒草甸种子雨的影响
2012-09-18文淑均李伟杜国祯
文淑均,李伟,2,杜国祯
(1.兰州大学生命科学学院,甘肃兰州730000;2.中国科学院水利部水土保持研究所,陕西杨凌712100)
前植物生产层
围栏封育对青藏高原东缘高寒草甸种子雨的影响
文淑均1,李伟1,2,杜国祯1
(1.兰州大学生命科学学院,甘肃兰州730000;2.中国科学院水利部水土保持研究所,陕西杨凌712100)
围栏封育是一种常见的退化草地恢复措施,其改变了草地植物生长繁殖的小环境。在青藏高原东缘高寒草甸中,采用定点监测的方法研究了封育8年之后围栏内外植被与种子雨的差异。结果表明,围栏封育显著降低了植被中物种丰富度、Shannon-Wiener指数以及植株个体密度,但显著提高了种子雨中物种丰富度和种子雨密度,种子雨密度增加主要由禾本科植物种子数量增加所致;种子雨中物种多度等级分布模式在封育地中符合生态位优先占据模型,而在放牧地中符合对数正态模型。围栏封育对种子雨的季节动态没有明显影响,表现为放牧地与封育地的种子雨均为单峰模式。封育地中种子雨与植被相似性略高于放牧地,但封育地与放牧地中种子雨具有极高相似性。
种子雨;封育;高寒草甸;青藏高原
种子雨是指在特定时间和特定空间从母株上散落一定数量的种子[1],是植物生命史动态过程中一个不可缺少的环节,更是预测土壤种子库的物种组成和大小以及植被更新演替趋势的有效手段。
高寒草甸是青藏高原地貌和气候等因素综合影响下形成的独有的地带性植被类型,主要分布于青藏高原海拔3 000~5 200m的地带,约占青藏高原1.4×108hm2天然草地的50%[2];由于过度放牧和气候变化,面临着不同程度的退化。围栏封育主要是通过人为措施降低或完全排除牲畜对草地生态系统的影响,使系统在自身的弹性下得以恢复和重建,是一种有效而且简便易行的促进草地恢复的策略[3],在青藏高原退化高寒草甸恢复中也被广泛应用。目前,封育对高寒草甸影响的研究主要集中在群落结构、物种多样性以及土壤理化性质方面[4-6],而围栏封育对种子雨的影响的研究相对不足,限制了深入解析围栏封育对草地恢复机理的诠释。因此,本研究分析封育对种子雨的物种组成、数量、多度等级以及时间动态的影响,明确种子雨与植被的关系,以期为退化草地的补播措施及准确预测退化草甸在长期禁牧之后的演替方向提供理论依据。
1 研究方法
1.1 研究区自然概况 本研究在兰州大学高寒草甸与湿地生态系统定位研究站进行,该站位于青藏高原东缘的甘肃省玛曲县境内,在县城以西7km处,其地理坐标为33°58′N,101°53′E,海拔3 500 m左右;年均温1.2℃,1月均温-10℃,7月均温11.7℃。年降水量约620mm,属高寒湿润气候;年日照时数约2 580h,年平均霜日大于270d。主体土壤类型为亚高山草甸土,以莎草科嵩草属(Kobresia),禾本科的羊茅属(Festuca)、早熟禾属(Poa)、剪股颖属(Agrostis)、披碱草属(Elymuns),菊科风毛菊属(Saussurea)以及毛茛科的银莲花属(Anemone)、毛茛属(Ranunculus)为优势植被类型,并伴生其他杂类草。从植物区系组成和水热特征来看,该区属典型的高寒草甸。封育地自1999年10月开始围栏封育,夏季禁牧,冬季少量放牧;放牧地与封育地毗邻,生长季正常放牧。
1.2 种子收集器的布置 于2007年6月中旬(绝大多数物种种子成熟之前),在封育地与放牧地中分别选取一块植被与微地形较为均一的样地。为了避免边缘效应,样地边缘与所处位置的边缘距离不少于30m;每块样地中设置4条28.5m的平行样线,样线间距为5m;每条样线上均匀放置20个15cm×15cm收集器(Astro Turf,美国生产),种子收集器间隔为1.5m;种子收集器放置之前用剪刀去除地表植物,用不锈钢钉将其固定于地表。分别于2007年7月30日、8月20日、9月10日、9月30日、10月20日(积雪覆盖之前),以收集器为单位分别收集各收集器上的种子,并将其装入信封带回实验室风干,在鉴定前保持种子干燥。
1.3 植被调查 于2007年8月上旬(处于生长季高峰期,能确保所有物种鉴定)进行植被调查。在每块样地沿对角线随机放置10个50cm×50cm的样方框,登记样方框内所有物种,并测量其高度、盖度和多度。
1.5 数据分析 用S∅rensen’s Similarity Coeffi-cient分别计算封育地和放牧地中种子雨与植被的相似性,用Shannon-Wiener指数(H′)分别计算植被和种子雨中的物种多样性。
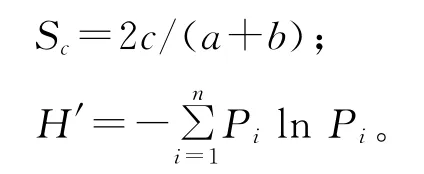
式中,Sc为相似系数,c为种子雨与植被共同物种数,a为种子雨中物种数,b为植被中物种数,Pi指第i个物种所占比例。
物种丰富度在植被中指单个样方内的物种数,在种子雨中指单个收集器内的物种数。种子雨密度换算成单位面积内种子数(粒·m-2)。
采用独立样本t检验分别对封育地与放牧地的植被和种子雨中的物种丰富度、Shannon-Wiener指数进行分析;为满足方差齐性,种子雨密度分析之前进行了自然对数转化。所有分析均在SPSS 16.0中完成。
为了验证封育对种子雨中物种多度等级分布模式的影响,在R软件vegan包中进行多度等级分布拟合[7]。各个模型解释如下。
Null拟合折棒模型(Broken Stick Model),其等级为r的物种多度期望值(ar)为:

式中,J为群落中个体数之和,S为群落中物种总数,x为从r到S中取值。
3)环境影响加权。根据各种环境影响的重要性级别,对标准化后的环境影响潜值进行赋权,从而评价其相对影响潜值大小。则
Preempt拟合生态位优先占据模型(The Niche Preemption Model),其等级r物种多度期望值为:

其唯一估计参数为优先占据系数α,即多度沿等级的衰减速率。
Lognormal拟合对数正态模型(log-Normal Model),它假定多度取对数后呈正态分布,其等级r物种多度期望值为:
式中,N为正态离差,M为随机变量的均值,σ为随机变量的方差。
Zipf拟合Z模型:
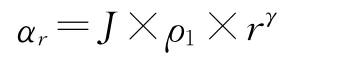
式中,ρ1为最多物种的拟合比例,γ为衰减系数。Zipfbrot是在Zipf模型中增加了一个参数,

Zipf模型中ρ1变成常数c,β为非线性参数。
封育地和放牧地的种子雨与植被物种组成相似性用无度量多维度标定排序(Nonmetric Multidimensional Scaling,NMDS)进行验证。NMDS是一种能在多维度空间展现相似矩阵并能维持原有项目之间关系等级的非参数排序技术[8],最终NMDS轴值表示有相似物种的点在排序空间越近,具体参考文献[9]。
2 结果
2.1 封育对植被的影响 在封育地与放牧地中总共调查出45个物种,共同种为40种,只在封育地与放牧地出现的物种分别为2和3种。均值比较结果表明,8年封育显著降低了物种丰富度、Shannon-Wiener指数以及多度(表1);NMDS排序结果显示,封育地与放牧地植被组成无明显分离(图1)。
2.2 封育对种子雨的影响 在封育地收集到4 581粒种子,分属于30种,13科;在围栏外收集到2 897粒种子,分属于31种,13科。封育地种子雨密度变化在711.11~9 111.11粒·m-2,平均2 545粒·m-2;放牧地种子雨密度变化在444.44~5 600粒·m-2,平均1 609.4粒·m-2。封育8年对种子雨中物种丰富度及种子雨密度均有显著影响,对Shannon-Wiener指数影响不显著(表2)。封育地中种子数量的增加主要由禾本科的披碱草、早熟禾、羊茅、剪股颖,毛茛科的小花草玉梅,伞形科的柴胡及玄参科的毛果婆婆纳所引起(图2)。

表1 封育对地上植被的影响Table 1 Effects of enclosure on vegetation
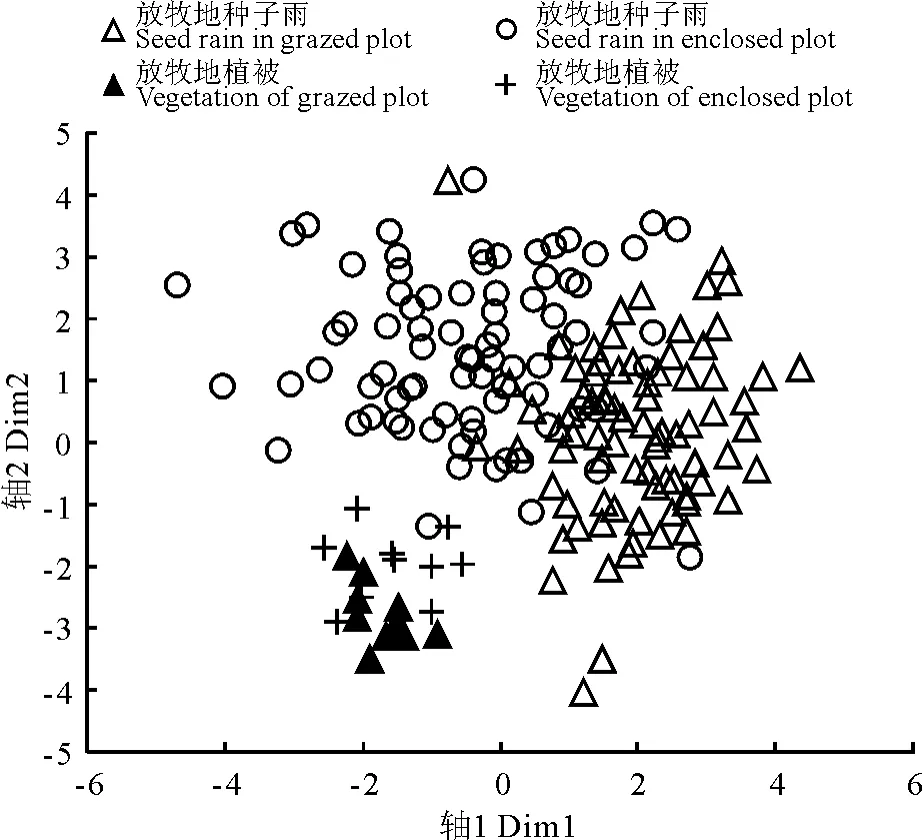
图1 种子雨与地上植被NMDS排序Fig.1 The NMDS ordination of vegetation and seed rain in different plots
2.3 封育对种子雨多度等级分布的影响 根据最小信息准则(Akaike Information Criterion,AIC)判断,封育对种子雨中物种多度等级分布模式产生了影响。在封育地中符合生态位优先占据模型,而放牧地中符合正态分布模型(表3、图3和图4)。
2.4 种子雨密度季节动态 在整个研究持续期,封育地与放牧地中的种子雨密度季节动态均呈单峰模式;封育地中由7月30日的125粒·m-2到9月30日逐渐上升为1 028.3粒·m-2,而10月20日急剧下降到610粒·m-2;放牧地中从7月30日的130粒·m-2到9月30日逐渐上升为753.3粒·m-2,10月20日急剧下降到146粒·m-2(图5)。

表2 封育对种子雨的影响Table 2 Effects of enclosure on seed rain
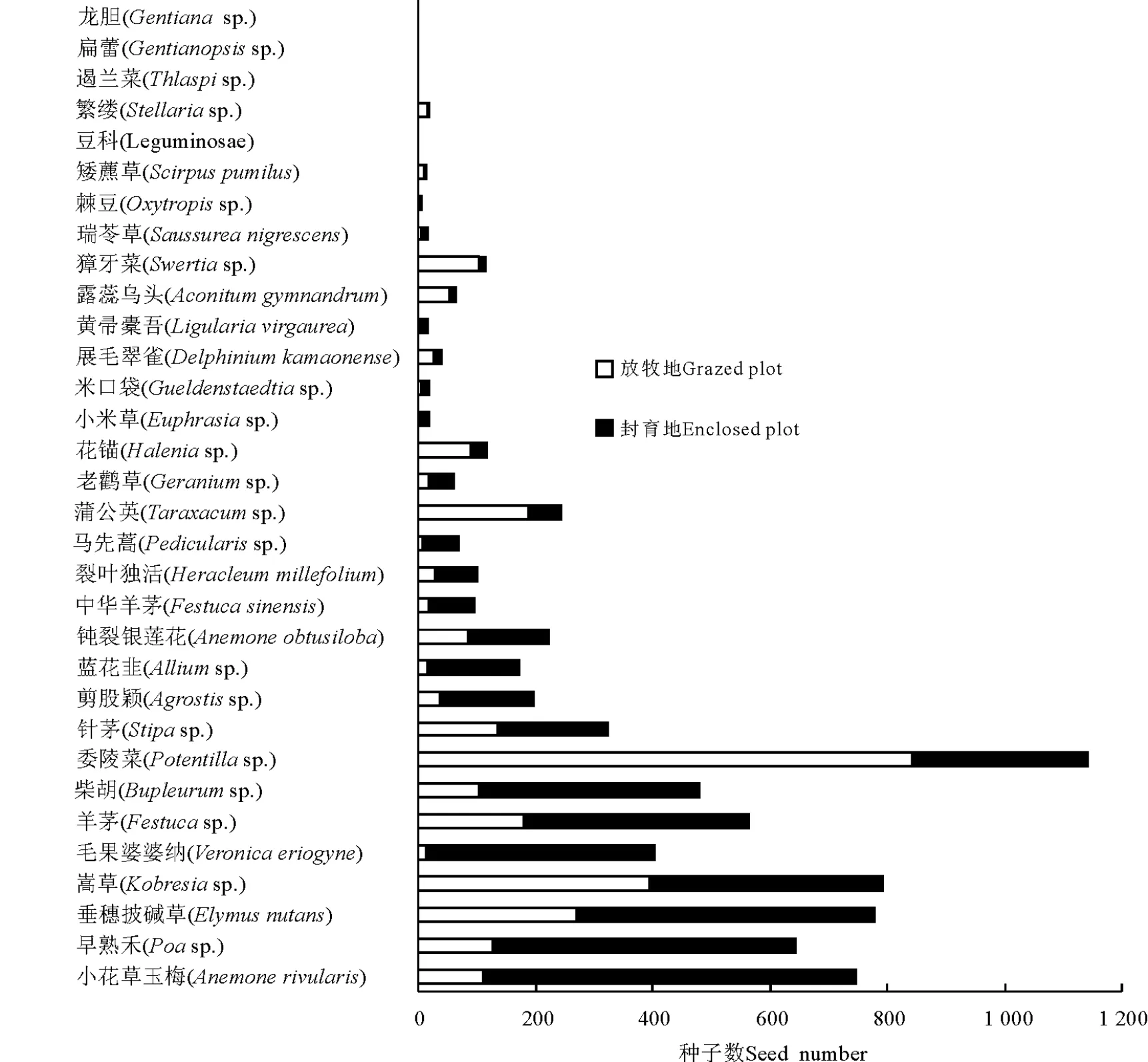
图2 放牧地与封育地中各物种种子数对比Fig.2 Comparison of seed number for each species

表3 不同模型拟合AIC值Table 3 AIC value for different models
2.5 种子雨与植被的关系 根据S∅rensen’s相似系数计算,在封育地和放牧地中种子雨与其植被物种组成相似性分别为0.667和0.595;而封育地与放牧地种子雨相似性高达0.951;封育地与放牧地植被相似性高达0.941(表4)。NMDS结果表明,种子雨与植被物种组成沿轴2有一定分离,但分离趋势并不明显;而封育地与放牧地中种子雨物种组成近乎没有分离,封育地与放牧地植被物种组成也近乎没有分离(图1)。
3 讨论
3.1 封育之后地表植被的变化 围栏封育能够增加退化草地物种丰富度和多样性[3],但也有长时间封育能降低物种丰富度、多样性指数以及植株个体密度的报道[4-6],这可能与封育时间的不一致有关。本研究中,封育导致植株个体密度的显著下降可能使得竞争力较强的物种(如禾本科植物)形成高而大的分蘖,从而抑制其他矮小植物的光吸收;或者是个体数较少的物种因个体消失而引起物种丰富度下降。这可以从光竞争假说[10]或者密度效应[11]得到理论支持。
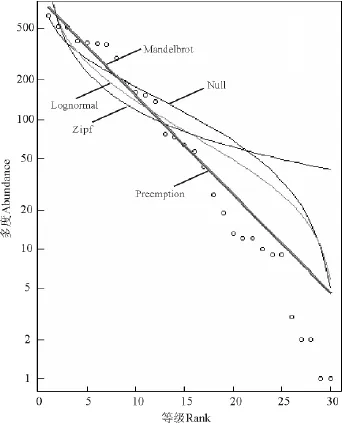
图3 封育地中种子雨多度等级Fig.3 Rank-abundance distribution of seed rain in enclosed plot

图4 放牧地中种子雨多度等级Fig.4 Rank-abundance distribution of seed rain in grazed plot
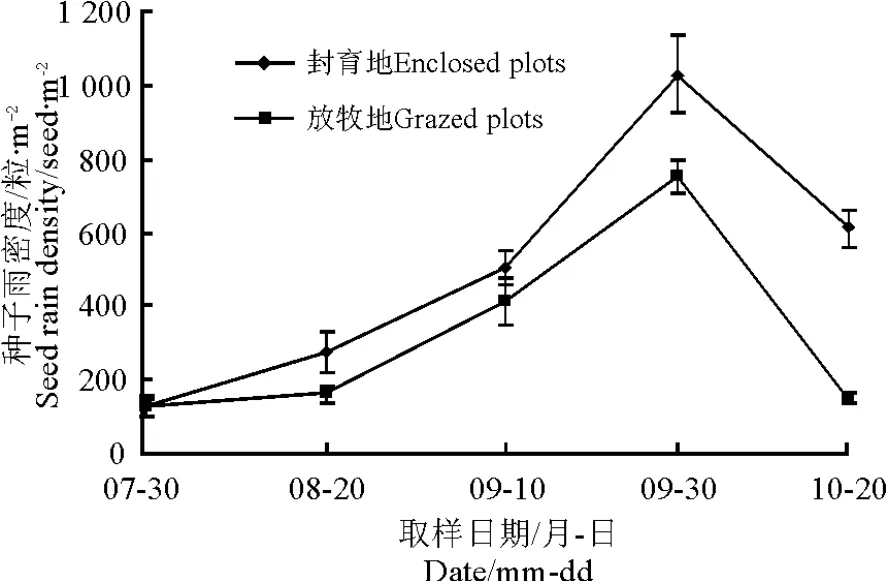
图5 种子雨密度季节动态Fig.5 The seasonal dynamics of seed rain density
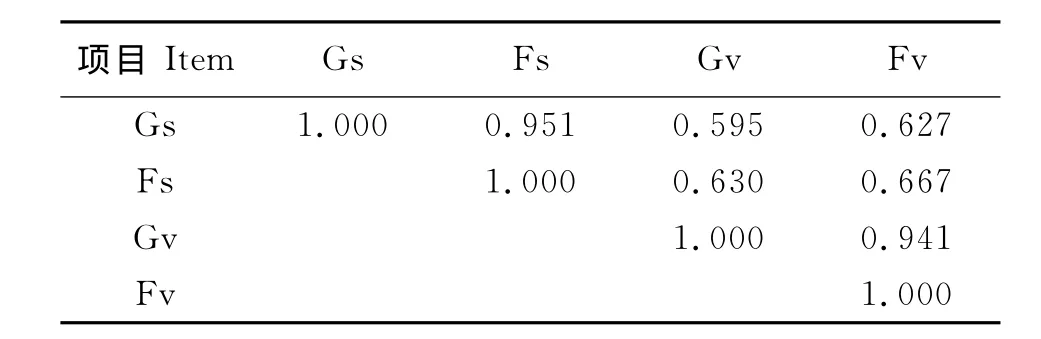
表4 种子雨与植被之间的相似系数Table 4 The S∅rensen’s similarity coefficient between seed rain and vegetation
3.2 封育之后种子雨的变化 封育因排除了动物采食,种子雨密度显著增加,这与Page和Harrington[12]的研究结果一致。封育对种子雨物种丰富度的影响与植被相反,更进一步支持了封育使优势种的优势度增加进而影响了其他非优势种的共存。本研究中,种子雨密度的增加主要依赖于禾本科的披碱草、早熟禾、羊茅、剪股颖和毛茛科小花草玉梅种子的增加,这可能由于禾本科植物属于优良牧草,在放牧地中会被牲畜选择性的采食;而小花草玉梅则是典型的毒杂草,在封育地中可避免家畜的践踏。
本研究表明,封育地与放牧地种子雨物种组成高度相似,但各物种种子数量差异显著,于是对比了封育地与放牧地中种子雨中物种多度等级分布模式,发现封育地中符合生态位优先占据模型,而放牧地中符合对数正态分布模型。生态位优先占据假说认为,第1位优势种占领生态位空间大部分,第2位占据余下的大部分,依次类推,末位的只能占有很少的空间;而对数-正态假说则认为物种对生态位的占有情形受诸如微气候、食物资源、空间资源等影响,因而物种分布呈现对数正态模式[13]。因种子雨对群落的潜在贡献,可以预测,长期封育会导致潜在群落向单顶级群落方向发展,这不利于生物多样性的保护与维持。
将不同类型草地中群落水平种子雨的密度[14-25](表5)与本研究结果进行对比,发现仅有高草草原(美国密西西比)、高寒草甸(中国)和白垩草地(法国)种子雨密度高于本研究。高草草原主要物种为禾本科、菊科、豆科和蔷薇科[14],白垩草地植物组成包含了由矮草地向高草地或灌丛次生演替阶段的物种[16],这些物种种子产量高并且海拔低,因而种子雨密度高于本研究。与邓自发等[15]的研究相比,对于同属于青藏高原地区,本研究样地种子雨密度明显偏低。分析本研究样地2003和2007年降水量,分别为731.4和612.9mm,可能降水减少尤其是返青季降水量不足影响了当年种子产量,导致种子雨密度偏低。相较于其他地区[17-25],本研究样地种子雨密度较大,封育样地内土壤种子库密度为3 883粒·m-2[26],高于本年度种子雨密度,可能原因有,种子库的形成是逐年种子雨累积形成的结果,且当地植被以多年生草本植物为主,幼苗更新更多依赖于营养繁殖,种子库中部分种子处于休眠状态;因此种子雨密度低于土壤种子库密度。
不同的研究表明,种子雨具有明显的季节动态[17,19,23],一般可以划分为起始、高峰、末期3个阶段。起始期是种子雨强度逐渐上升的阶段,种子雨开始时先在局部发生,逐渐到全面发生,并达到一定密度;高峰期是指种子雨强度上升到最大并持续一段时间后,然后缓慢下降,这段时期为高峰期;末期是指种子的分布接近尾声种子雨强度已经很小,并接近为0,种子的初次分布趋于结束。本研究中围栏内外种子雨的模式与之基本一致,因该研究点冬天积雪覆盖,种子收集期没有持续到种子散布的末期。

表5 不同类型草地中种子雨对比Table 5 The comparison of seed rain in different habitats
3.3 种子雨与植被关系 围栏内外种子雨与植被相似性分别为0.667和0.595,英格兰不同的撂荒白垩矿旧址中种子雨与植被相似性为0.64和0.72[22],美国衣阿华自然草原湿地与恢复湿地中种子雨与植被相似性分别为0.53和0.50[17]。这些都说明种子雨与植被有很强的相关性,种子雨物种的组成很大程度依赖于植被的物种组成。对比本研究的种子库,封育地中种子库与植被相似性仅为0.453[25],明显低于种子雨与植被的相似性系数,可能因为一部分种子在散落后未来得及形成种子库已被动物采食或者腐烂,也可能是种子鉴别困难而将相近物种归于同一属导致了相似性被高估。
尽管封育对地上植被和种子雨中的物种丰富度及种子雨密度影响显著,但围栏内外种子雨相似性高达0.951。进一步说明围栏封育对群落的影响主要体现在优势种的改变,对于物种组成影响有限,该结论也得到了NMDS排序结果的支持。
4 结论
长时间的封育虽然增加了种子雨中的物种丰富度和种子雨密度,但降低了植被中的物种丰富度和个体密度;这也是牧草产量增加与生物多样性保护中的难题[4]。当前草地恢复中补播措施一般选用的禾本科植物种子[27],本研究表明,封育后种子雨密度增加主要是禾本科植物种子数增加所引起,在同类高寒草甸中植被恢复至少不受禾本科种子所限制;补播种的选择应更为广泛。
[1]Harper J L.Population Biology of Plant[M].London:Academic Press,1977.
[2]石福孙,吴宁,罗鹏,等.围栏禁牧对川西北亚高山高寒草甸群落结构的影响[J].应用与环境生物学报,2007,13(6):767-770.
[3]左万庆,王玉辉,王风玉,等.围栏封育措施对退化羊草草原植物群落特征影响研究[J].草业学报,2009,18(3):12-19.
[4]Wu G L,Du G Z,Liu Z H,et al.Effect of fencing and grazing on a Kobresia-dominated meadow in the Qinghai-Tibetan Plateau[J].Plant and Soil,2009,319: 115-126.
[5]刘德梅,马玉寿,张德罡,等.封育对“黑土滩”垂穗披碱草栽培草地群落结构和特征的影响[J].草业科学,2009,26(10):59-66.
[6]李慧,蒋平安,程路明,等.围栏对新疆山区草地植物群落多样性的影响[J].新疆农业大学学报,2005,28(3):40-43.
[7]Jari O F,Guillaume B,Roeland K,et al.vegan:Community Ecology Package.R package version 2.0-0[EB/OL].http://CRAN.R-project.org/package=vegan 2011.
[8]Legendre P,Legendre L.Numerical Ecology[M].New York:Elsevier,1998.
[9]Ma M J,Zhou X H,Du G Z.Soil seed bank dynamics in alpine wetland succession on the Tibetan Plateau[J]. Plant and Soil,2011,346(1):19-28.
[10]Schippers P,Kropff M J.Competition for light and nitrogen among grassland species:A simulation analysis[J].Functional Ecology,2001,15(2):155-164.
[11]Stevens M H H,Carson W P.Plant density determines species richness along an experimental fertility gradient[J].Ecology,1999,80(2):455-465.
[12]Page M J,Harrington R A.Fourteen months of seed rain in three Australian semi-arid communities[J]. Austral Ecology,2009,34(3):294-305.
[13]孙儒泳.动物生态学原理[M].北京:北京师范大学出版社,2001.
[14]Rabinowitz D,Rapp J K.Seed rain in a North American tallgrass prairie[J].Journal of Applied Ecology,1980,17(3):793-802.
[15]邓自发,谢晓玲,王启基,等.高寒小嵩草草甸种子库和种子雨动态分析[J].应用与环境生物学报,2003,9(1):7-10.
[16]Chabrerie O,Alard D.Comparison of three seed trap types in a chalk grassland:Toward a standardised protocol[J].Plant Ecology,2005,176(1):101-112.
[17]Spence J R.Seed rain in grassland,herbfield,snowbank,and fellfield in the alpine zone,Craigieburn Range,South Island,New Zealand[J].New Zealand Journal of Botany,1990,28(4):439-450.
[18]Kettenring K,Galatowitsch S.Seed rain of restored and natural prairie wetlands.[J].Wetlands,2011,31(2):283-294.
[19]Urbanska K M,Fattorini M.Seed rain in high-altitude restoration plots in Switzerland[J].Restoration Ecology,2000,8(1):74-79.
[20]Welling P,Laine K,Bakker J P.Regeneration by seeds in alpine meadow and heath vegetation in sub-arctic Finland[J].Journal of Vegetation Science,2002,13:217-226.
[21]Urbanska K M,Fattorini M.Seed rain in natural grassland and adjacent Ski Run in the Swiss Alps:A preliminary report[J].Restoration Ecology,1998,6(2):159-165.
[22]Chacón P,Cavieres L A.Recovery of alpine plants after a severe human disturbance in the Andes of central Chile[J].Arctic,Antarctic and Alpine Research,2008,40:617-623.
[23]Jefferson R G,Usher M B.Seed rain dynamics in disused chalk quarries in the Yorkshire Wolds,England, with special reference to nature conservation[J].Biological Conservation,1989,47(2):123-136.
[24]Molau U,Larsson E L.Seed rain and seed bank along an alpine altitudinal gradient in Swedish Lapland.[J]Canadian Journal of Botany,2000,78:728-747.
[25]Larsson E L.Two ways of trapping seeds in alpine environments,Lapland,Sweden[J].Nordic Journal of Botany,2003,23(3):333-343.
[26]马妙君,周显辉,吕正文,等.青藏高原东缘封育和退化高寒草甸种子库差异[J].生态学报,2009,29(7):3658-3664.
[27]郑华平,陈子萱,牛俊义,等.补播禾草对玛曲高寒沙化草地植物多样性和生产力的影响[J].草业学报,2009,18(3):28-33.
Effects of enclosure on seed rain of alpine meadow in the eastern Tibetan Plateau
WEN Shu-jun1,LI Wei1,2,DU Guo-zhen1
(1.School of Life Sciences,Lanzhou University,Lanzhou 730000,China 2.Institute of Soil and Water Conservation of Chinese Academy of Sciences and Ministry of Water Resources,Yangling 712100,China)
Enclosure has been widely used in degraded grassland restoration as a brief and effective treatment,which changed micro-conditions of growth and production for plants.In the present study,the vegetation and the seed rain were investigated in enclosed and grazed plots after 8-year enclosing on alpine meadow in the eastern Tibetan Plateau.The results showed that enclosure has significantly negative effects on species richness,Shannon index and individual density of vegetation,however,enclosure has significantly positive effects on richness and density of seed rain in enclosed plots compared with those in grazed plots.The changes of the numbers of Gramineae seeds played a key role in increasing of seed rain density.RAD(rank abundance distribution)displayed different patterns from enclosed plots to grazed ones.In enclosed and grazed plots,the pattern for seasonal dynamics of seed rain was unimodal.The S∅rensen’s similarity coefficient between seed rain and vegetation at fenced plots was higher than that at grazed plots,but the S∅rensen’s similarity coefficient between seed rain in enclosed plots and that in grazed plots was very high.
seed rain;enclosure;alpine meadow;Tibetan Plateau
DU Guo-zhen E-mail:guozdu@lzu.edu.cn
S812.2
A
1001-0629(2012)03-0333-08
2011-11-30 接受日期:2012-01-09
国家科技支撑项目(2009BAC53B01)
文淑均(1979-),男,湖北松滋人,在读博士生,研究方向为群落生态学。E-mail:wenshujun@gmail.com
杜国祯 E-mail:guozdu@lzu.edu.cn
