结缕草属植物部分外部性状的遗传分析
2012-08-20郭海林陈宣薛丹丹刘建秀
郭海林,陈宣,薛丹丹,刘建秀
(江苏省中国科学院植物研究所 南京中山植物园,江苏 南京210014)
结缕草属(Zoysia)植物是禾本科画眉草亚科的多年生草本植物,是优良的暖季型草坪草。可广泛应用于观赏草坪、运动草坪、休憩草坪以及庭院草坪等各种草坪的建设中[1-4]。
对结缕草属植物的深入研究有利于培育出优良的结缕草属植物新品种。研究表明,结缕草属植物大约有11种和一些变种与变型[1],结缕草属植物的开花习性为雌蕊先熟雄蕊后熟,是一种典型的常异花授粉植物[5],结缕草属植物的染色体倍性为2n=4X=40[6,7]。结缕草属植物不同种、同一种的不同种质资源间在外部性状[8,9]、抗性(抗寒、旱、盐、病等)等方面[10-16]均存在着丰富的遗传变异,这些研究均为结缕草属植物新品种选育奠定了良好的基础。其中外部性状的表现与研究直接关系到草坪草坪用质量的好坏与改良。而外部性状的遗传机制对其遗传改良和选择适当的育种方法均具有重要的指导意义,若这些性状是由1~2个主基因控制则可采用单交重组或简单回交转育的方法;若这些性状由多个微效基因控制,则因为涉及的基因多、单个基因效应小、增效基因集中到一个个体的概率低、从表型鉴别基因型的难度大等特点,则要采用聚合回交(或双回交)及轮回选择累积增效基因的方法[17]。
盖钧镒等[17]提出了植物数量性状的主基因-多基因遗传理论,认为多数性状由主基因加多基因共同控制是数量性状遗传的基本模型,而纯主基因或纯多基因的遗传模型只是特例,并建立了植物数量性状主基因+多基因混合遗传模型分析方法[16-18],该模型自开发以来,已在小麦(Triticum aestivum)[19,20]、水稻(Oryza sativa)[21]、大豆(Glycine max)[22]、油菜(Brassica campestris)[23]、草坪[24]等多种植物的遗传分析中得到了广泛的应用,在这些植物的遗传机制研究方面发挥了重要作用。受上述思想的启迪,本研究试图应用主基因+多基因混合遗传模型分析方法对2份结缕草属种源的正反交F1分离群体的8个与坪用价值相关的营养性状进行遗传分析,以明确结缕草属植物营养性状的遗传特性,为结缕草属植物品种改良、相关性状的分子标记、定位以及标记辅助育种等方面提供科学依据。
1 材料与方法
1.1 供试材料
供试材料为结缕草(Zoysiajaponica)‘J36’和中华结缕草(Zoysiasinica)Z039相互杂交的168个正交后代和99个反交后代,这些后代已通过SRAP分子标记技术鉴定为真杂种。亲本‘J36’为从美国引进的结缕草品种,Z039为从安徽宁国采集的种源材料,两亲本在外部形态上存在较大的差异,两亲本来源及外部性状的观测值见表1。两亲本均种植于江苏省中科院植物研究所的苗圃地内,287个杂交后代的获得及其培养情况如下:2006年4月上旬通过控制授粉法对外部性状存在显著差异的2份亲本材料J36和Z039进行相互杂交,结果成功地获得了184个正交后代和103个反交后代,并于2006年7月上旬将杂交后代种子用30%的NaOH处理80min后首先在培养皿培养,待第1片叶长到1.5~2.0cm时将其移入穴盘,其中穴盘的每一穴位只移入1株。2007年4月将后代从穴盘移入口径为22cm的塑料盆中,每盆1株。对杂交后代进行常规管理,2007年7-9月,对杂交后代通过SRAP分子标记技术进行杂种真实性鉴定,结果有168个正交后代和99个反交后代被鉴定为真杂种,这些真杂种用于对本研究的结缕草属植物外部性状的遗传分析。
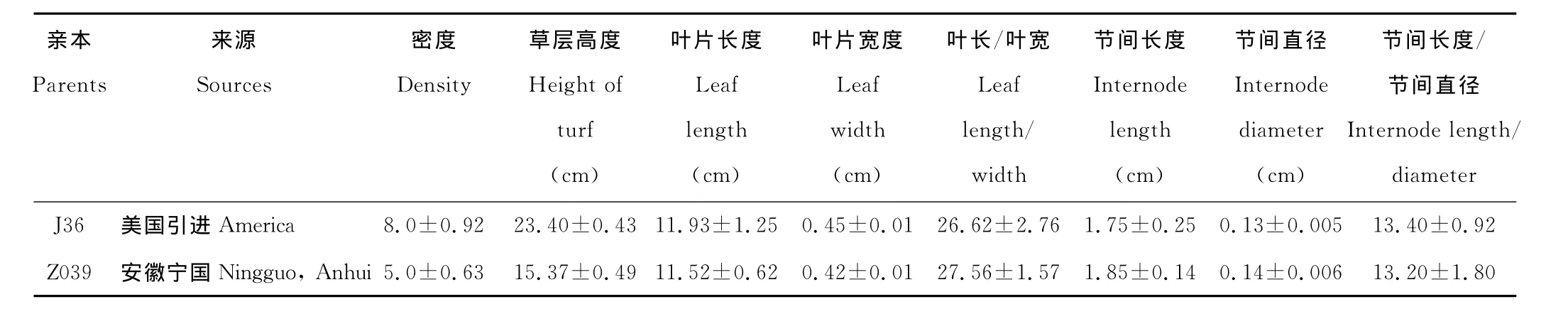
表1 亲本来源及其外部性状观测值Table 1 Sources and the characteristics of morphological characters of parents
1.2 外部性状观测
外部性状的观测于2008年7-8月份进行,观测指标包括密度、草层高度、叶长与叶宽、叶片长宽比、匍匐茎长度和直径和匍匐茎长度与直径比。观测方法如下:
密度:具体指草层中直立枝的密度。观测采用9级制打分法,其中1为密度最低、6为可以接受,9为密度最高,为3个观测值的平均值。
草层高度:指结缕草生长的自然高度,采用三点法测量。叶长与叶宽:指成熟叶片的叶长和叶宽,重复10次。节间长度与直径:指第3节的长度和直径,重复10次。
1.3 数据分析
结缕草属植物为常异交植物,亲本材料存在高度的异质性,F1世代就产生性状分离现象,类似于纯合亲本杂交的F2群体。本研究的遗传数据分析应用植物数量性状主基因+多基因混合遗传模型,采用单个分离世代F2群体的数量性状分离分析方法[16-18]对正交J36×Z039及反交Z039×J36的F1群体的外部性状进行分析,由模型的似然函数值计算出AIC(Akaike’s information criterion)值,AIC值最小的模型为相对最佳模型,通过比较无主基因、1对主基因(A)和2对主基因(B)共11种遗传模型的AIC值,并进行遗传模型的适合性测验,包括均匀性检验(U12,U22和 U32),Smirnov检验(nW2)和Kolmogorov检验(Dn),综合考虑极大似然函数、AIC值以及适合性检验的结果,最终确定最优模型。在此基础上,估计最优模型的遗传参数,包括加性遗传效应(d)和主基因遗传率(h2mg),并通过Excel软件的数据分析功能,计算平均值、变异范围、变异系数、偏度系数、峰度系数和方差等指标。植物数量性状主基因+多基因遗传体系的分析统计软件由南京农业大学国家大豆改良中心提供。
2 结果与分析
2.1 F1群体的营养性状遗传变异
对J36×Z039的168份杂交后代和反交Z039×J36的99份杂交后代的外部性状观测分析,发现不同性状正反交的变异范围均较大(表2),所观测的8个性状中,在正交后代群体中变异系数由大到小为:密度(29.66%)>节间长度/直径(24.23%)>节间长度 (21.06%)>叶长(17.19%)>草层高度(17.03%)>节间直径(17.0%)>叶长/叶宽(16.98%)>叶宽(8.79%);反交后代群体中变异系数由大到小为:密度(26.44%)>节间长度/直径(22.72%)>节间长度 (18.14%)>叶长/叶宽(16.53%)>草层高度(16.39%)>节间直径(15.77%)>叶长(15.57%)>叶宽(8.31%),比较正反交后代的变异系数可以看出,在这8个性状中,不同后代间密度的变异最大,其次为节间长度/直径和节间长度,叶宽的变异最小,其他性状的变异居中。从变异范围来看,正反交杂交后代中每一个性状的变异范围均超出了双亲的变异范围,表明控制这些性状的增效和减效基因在双亲中呈分散分布,通过基因重组可产生正向和负向的超亲后代。
不同性状正反交后代的平均值略有差异(表2)。为比较正反交后代间性状表现的差异显著性,对不同性状的正反交后代观测值间进行了t-检验。t-检验结果表明(表2),所检测的8个性状中,只有密度和节间长度正反交后代间的t统计值小于t双尾临界值1.96(α=0.05),正反交后代间没有显著差异,其余6个性状草层高度、叶长、叶宽、叶长/叶宽、节间直径和节间长度/直径的正反交后代的t统计值绝对值均大于t双尾临界值1.96(α=0.05),正反交后代间存在显著差异,由此推测这6个性状可能存在母体遗传效应。
根据正反交F1群体8个营养性状的观测结果,对正反交的每个性状分别作次数分布图(图1)。不同性状均呈连续性分布,其中叶长(反交)(图1-C2)和叶宽(正交)(图1-D1)近似正态分布,草层高度(反交)(图1-B2)和叶长(正交)(图1-C1)近似于双峰分布,其他性状均呈偏态分布,符合主基因+多基因的遗传特征。
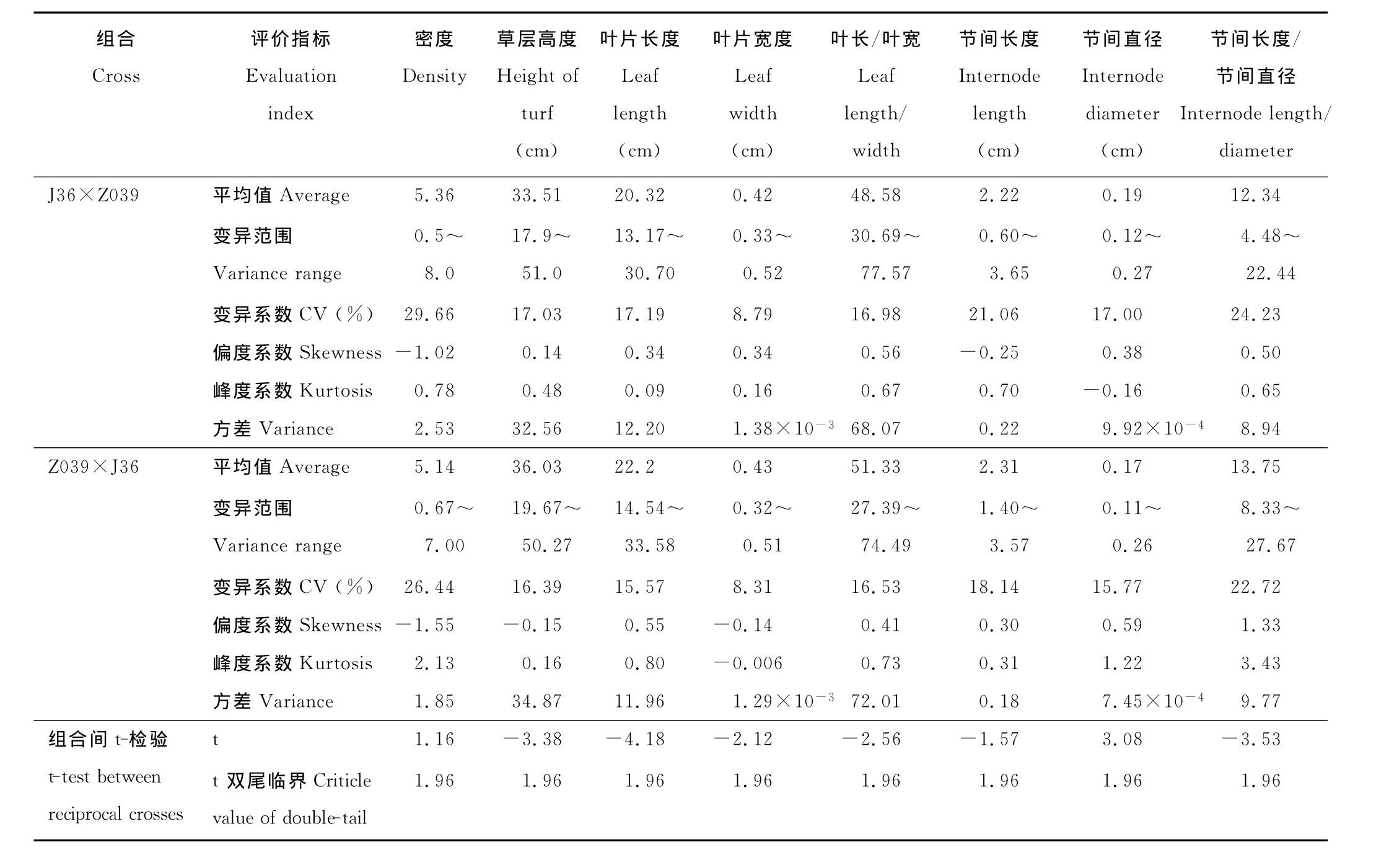
表2 J36×Z039正反交杂交后代营养性状的变异分析Table 2 The variation of vegetative characters of reciprocal progenies in J36×Z039
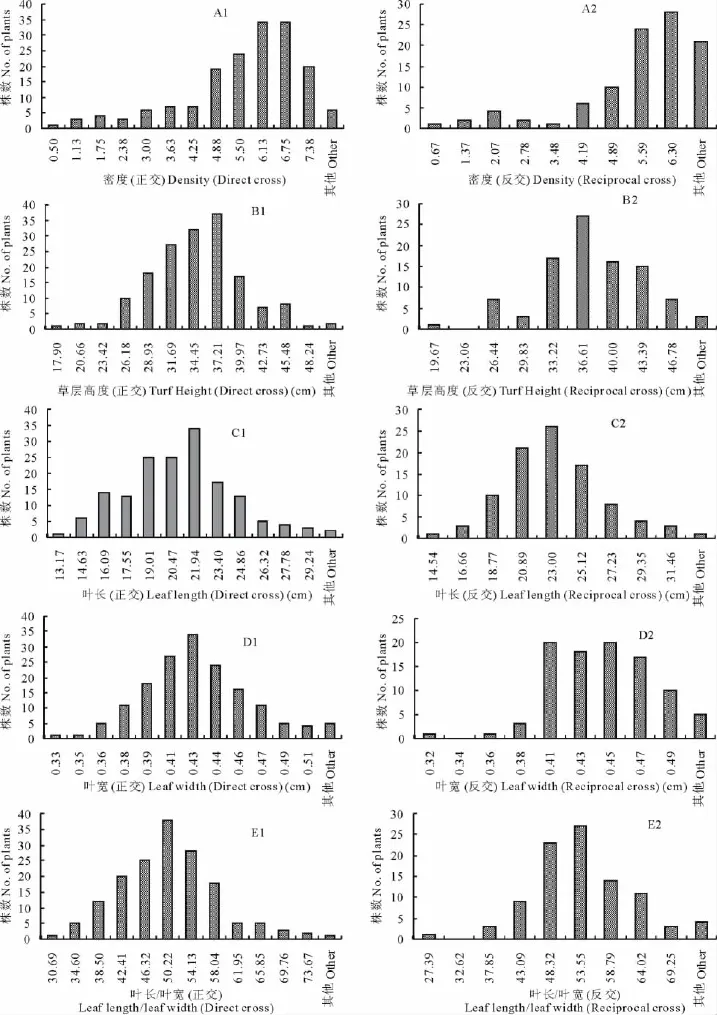
图1 J36×Z039正反交杂交后代群体8个营养性状的次数分布图Fig.1 Frequency distribution of 8vegetative characters of reciprocal F1populations of J36×Z039
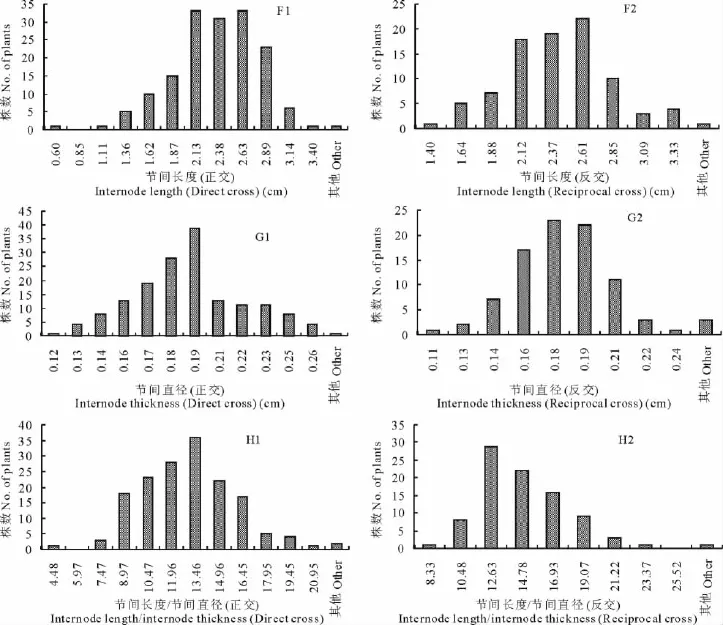
续图1 J36×Z039正反交杂交后代群体8个营养性状的次数分布图Fig.1 (continued) Frequency distribution of 8vegetative characters of reciprocal F1populations of J36×Z039
2.2 F1群体的营养性状主基因+多基因混合遗传分析
分别计算8个营养性状的正反交F1群体在11种遗传模型下的极大似然函数值和AIC值(表3),以AIC值较小的模型作为候选模型。结果表明,在被检验的各种模型中,密度正反交都是B-1模型的AIC值最小,适合性检验结果(表4)也表明,正反交F1群体中密度的分布与理论分布是一致的(P>0.05),因此,B-1模型,即“两对主基因,加性-显性-上位性模型”是J36×Z039正反交F1群体密度的最适遗传模型,其中正交的加性效应(da和db)分别为1.936 5和1.063 7,显性效应(ha和hb)分别为0.575 4和0.035 6,上位性效应(i,jab,jba,l)分别为-0.651 1,-0.187 2,-0.266 6和0.193 6,主基因遗传方差(σ2mg)为2.37,主基因遗传率[h2mg(%)]为93.67%,反交的加性效应为1.282 4和0.830 8,显性效应为0.842 8和0.490 7,上位性效应为-0.829 1,-0.490 2,-0.398 2和-0.086 5,主基因遗传方差为1.169 6,主基因遗传率为63.22%(表5)。
草层高度、叶片长度、叶片宽度、叶长/叶宽、节间长度5个性状正反交F1群体均表现为A-0模型的AIC值最小(表3),适合性检验结果发现,前3个性状与节间长度反交群体的分布均与理论分布一致(P>0.05),叶长/叶宽的正反交群体表现和节间长度的正交群体表现分别有一个值与理论值存在显著差异,而其他值均与理论分布一致,因此,认为A-0模型,即无主基因模型是这5个性状的最适遗传模型,也就是说在正反交F1群体中,这5个性状表现出由误差或误差+多基因所修饰的单一正态分布特征。
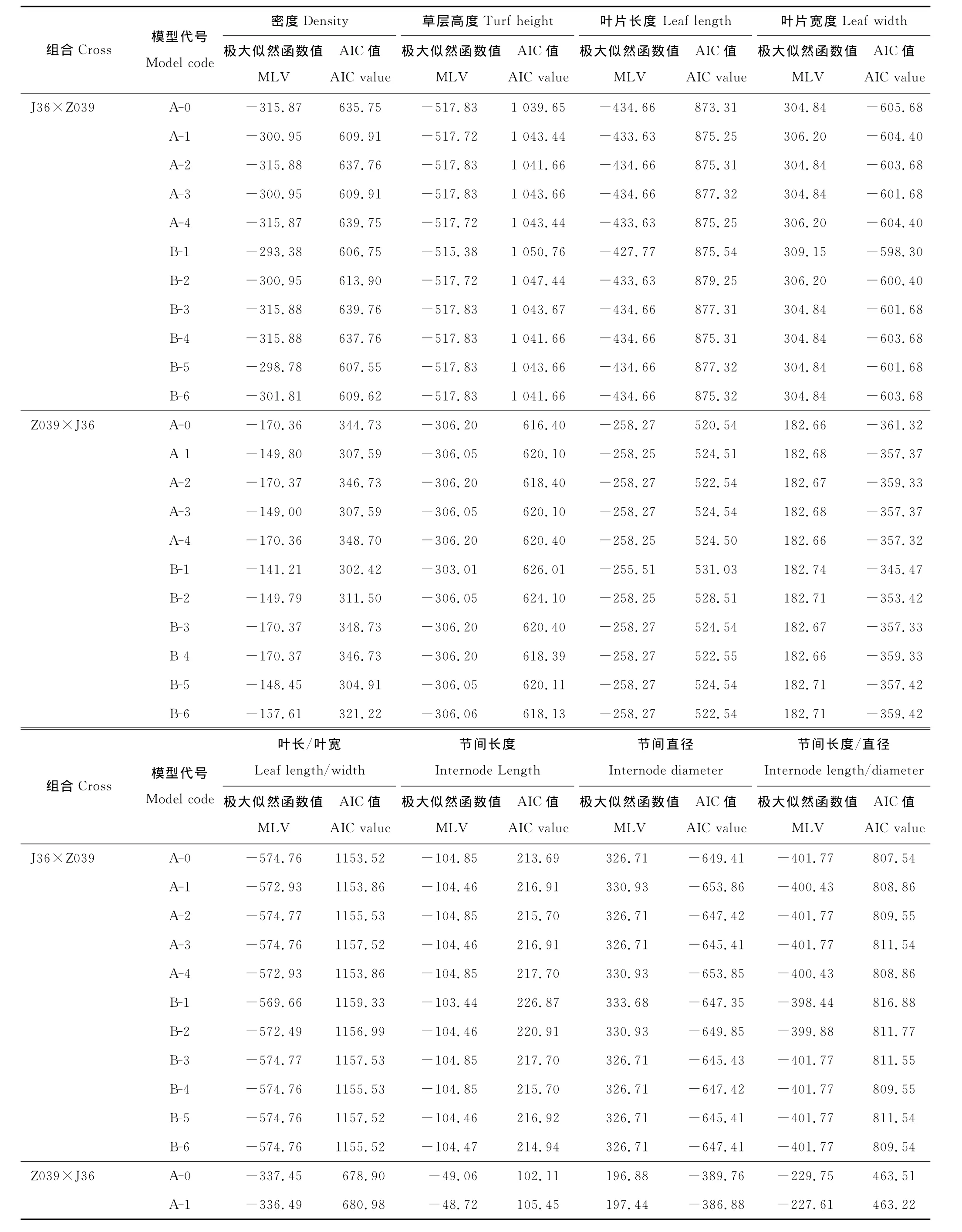
表3 不同遗传模型下的极大似然函数值和AIC值Table 3 Maximum likelihood value(MLV)and Akaike’s information criterion(AIC)under different genetic model
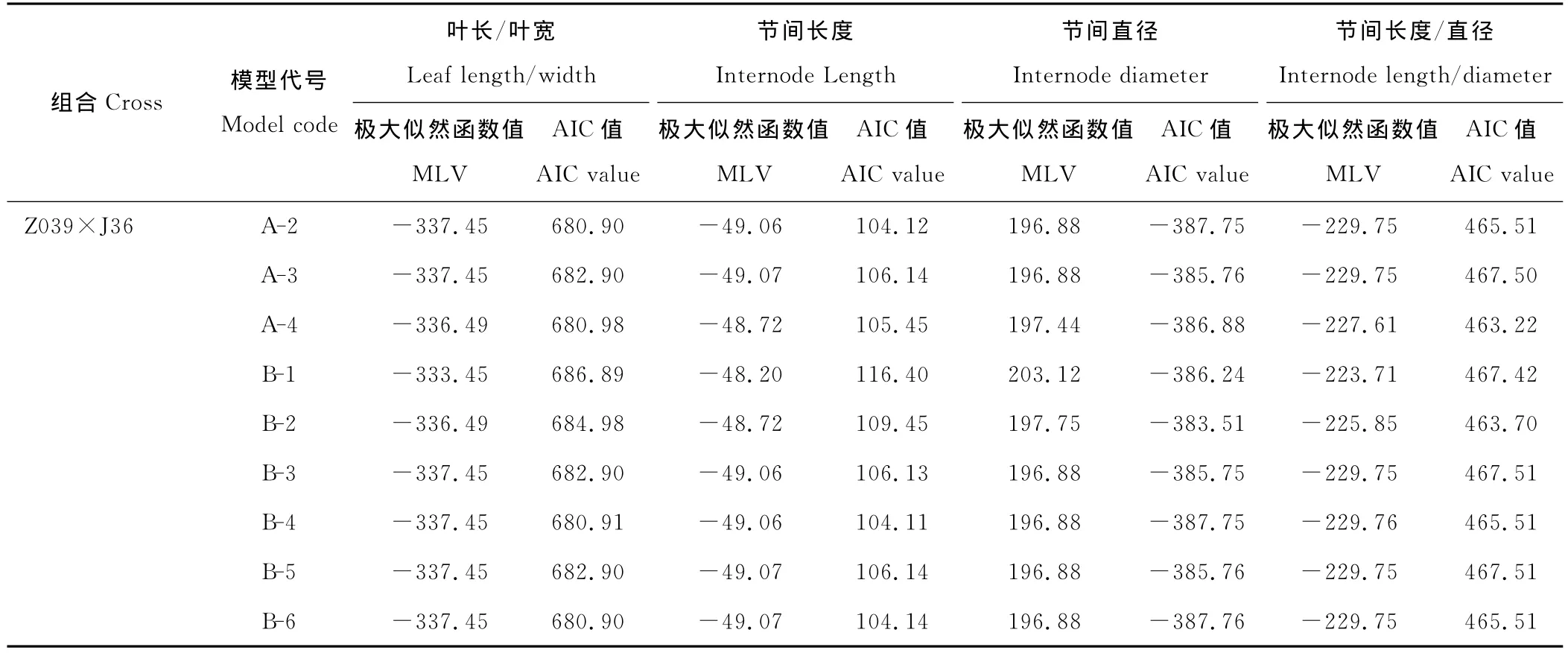
续表3 Continued
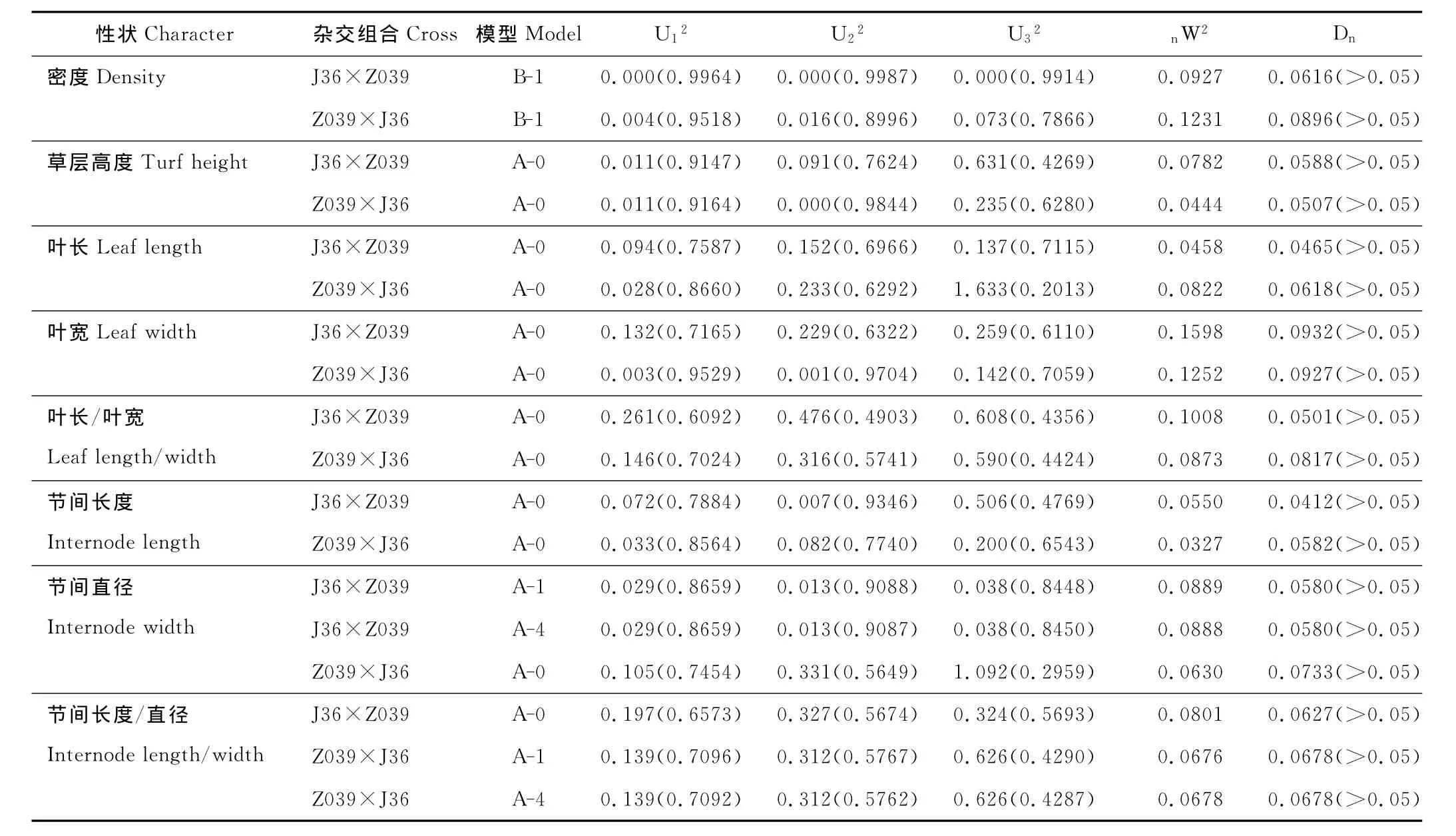
表4 入选模型的适合性检验Table 4 Tests for goodness-of-fit of selected model
节间直径正交后代群体检测结果为A-1和A-4模型的AIC值最小,最适模型为一对主基因控制的遗传模型,反交是A-0模型的AIC值最小,即无主基因模型为最适遗传模型;节间长度的检测结果为正交是A-0模型的AIC值最小,最适模型为无主基因模型,反交为A-1与A-4模型的AIC值最小,最适模型为一对主基因控制的模型。正反交的遗传模型不一致,这可能与节间直径受母体遗传效应影响的结果有关。

表5 B-1模型下F2群体密度一阶遗传参数的计算Table 5 Estimates of genetic parameters under genetic model B1for density of zoysia F2populations
3 结论与讨论
本研究将2份外部性状存在差异的结缕草与中华结缕草种源材料进行杂交,对获得的正反交F1后代的8个与坪用价值相关的外部性状的遗传变异进行了分析。从变异范围来看,正反交杂交后代中每一个性状的变异范围均超出了双亲的变异范围,表明控制这些性状的增效和减效基因在双亲中呈分散分布,通过基因重组可产生正向和负向的超亲后代。但从变异系数发现,不同性状通过杂交育种的手段可以改良的程度不同,综合分析正反交后代不同性状变异系数的统计结果表明,在这8个性状中,不同后代间密度的变异系数最大,其次为节间长度/直径和节间长度,叶宽的变异最小,其他性状的变异居中。变异系数反映了待改良群体的变异情况,变异越大,说明选择潜力越大,因此,在本研究中,通过性状的变异系数可以发现,密度和节间长度2个性状比较容易通过杂交育种加以改良,而叶片宽度的变异系数很低,表明叶宽很难通过杂交育种加以改良。刘建秀等[9]对中华结缕草种质资源的形态变异情况进行了研究,结果发现节间直径和叶宽在所观测性状中变异系数最小,分别为12.5%和14.4%,认为通过系统选育的方法对中华结缕草的叶宽和节间直径进行改良,其效果明显低于叶长和草层高度等性状。结合本研究的结果,认为结缕草和中华结缕草的叶宽变异较小,很难通过系统选育的手段对其进行改良。
对于纯合亲本来说,母体遗传效应的研究一般通过正反交F1的表型值与双亲进行比较来确定,而结缕草属植物的杂交F1代即发生分离,无法通过单个的F1数据来与亲本进行比较,因此,本研究通过t测验的方法,根据正反交后代群体间的差异显著性来推测不同性状是否有母体遗传效应,结果发现,8个性状中有6个性状正反交后代间有显著差异,初步推测这些性状可能存在母体遗传效应,这个结论还有待于在以后的研究中进一步加以验证。
本研究应用盖钧镒等[17]提出的主基因+多基因混合遗传模型对结缕草与中华结缕草杂交的F1群体的8个营养性状进行了遗传分析,通过比较AIC值确定了每个性状的最佳遗传模型,为这些性状遗传机制的进一步研究和遗传改良奠定了基础。遗传模型的研究结果发现,所研究的8个性状中只有密度为2对主基因的加性-显性-上位性遗传模型,而其他性状几乎全部为无主基因模型,节间直径的正交后代和节间长度/直径的反交后代表现为1对主基因的遗传模型,但是这2个性状对应的反交后代和正交后代也表现为无主基因模型,这一方面可能是受母性遗传的效应,同时也可能是受环境的影响,数量性状基因数目有多有少,各对基因效应的大小也不同,而且其表现受环境的影响,可能对某个性状的特定群体来说控制这个性状的基因在已有的环境条件下主基因效应明显,而在另外一个群体中在相应的环境条件下效应较弱,表现为微效基因,关于A-0模型的遗传机制,须通过同质群体提供误差方差以鉴别其有无遗传变异存在。
结缕草属植物为常异交植物,亲本异质性很高,因此在F1代即发生分离现象。本研究借助于纯合亲本杂交的F2群体的主基因+多基因混合遗传模型对结缕草属植物的8个营养性状进行了遗传分析,初步明确了正反交杂交后代的遗传模型,而实际上利用单个分离世代的分离分析方法是分两步骤进行的,首先鉴定主基因是否存在,然后再根据同质群体P1、P2和F1提供试验误差方差的无偏估计后鉴定是否有多基因的修饰。因模型的限制,本研究在只有单个亲本和F1群体的表型值的情况下,只对主基因的存在情况和主基因遗传效应与遗传率进行了检验,关于多基因的存在和效应都无法鉴别。
目前,关于农作物等自交植物,遗传背景高度纯合,容易得到自交群体的植物中,遗传学方面的研究已比较深入,针对不同的研究需要建立了很多生物学研究体系,而对于草坪草和大多数林木植物等常异交植物,因遗传背景高度杂合、遗传负荷大,给遗传学的研究带来了很大的困难,目前在遗传分析、遗传图谱构建等遗传学方面没有一套合适的生物学研究体系可以利用,因此要对上述植物的遗传机制进行更全面的分析,还有待于建立更完善的分析方法。
[1] Engelke M C,Anderson S.Zoysiagrasses(Zoysiaspp.)[A].In:Michael D C,Ronny R D.Turfgrass Biology,Genetics and Breeding[M].New Jersey:John Wiley and Sons,Inc.,2002:271-285.
[2] 董厚德,宫莉君.中国结缕草生态学及其资源开发与应用[M].北京:中国林业出版社,2001.
[3] 郭海林,刘建秀.结缕草属植物育种进展概述[J].草业学报,2004,13(3):106-112.
[4] 胡化广,刘建秀,宣继萍,等.结缕草属植物的抗旱性初步评价[J].草业学报,2007,16(1):47-51.
[5] 郭海林,刘建秀.三种暖季型草坪草的开花习性及自交结实率的研究[J].草原与草坪,2001,(增刊):45-47.
[6] Forbes I.Chromosome numbers and hybrids in zoysia[J].Agronomy Journal,1952,44:194-199.
[7] Yaneshita M,Kaneko S,Sasakuma T.Alleotetraploid of zoysia species with 2n=40based on a RFLP genetic map[J].Theoretical and Applied Genetics,1999,98(5):751-756.
[8] 李亚,凌萍萍,刘建秀.中国结缕草属植物(Zoysiaspp.)地上部分形态类型多样性[J].植物资源与环境学报,2002,11(4):33-39.
[9] 刘建秀,郭爱桂,郭海林.中华结缕草种质资源形态变异及其形态类型[J].草地学报,2003,11(3):189-196.
[10] 李亚,谢晓金,宣继萍.中国结缕草属(Zoysiaspp.)植物抗寒性评价[J].草地学报,2003,11(3):240-245.
[11] 胡化广,刘建秀,宣继萍,等.结缕草属植物的抗旱性初步评价[J].草业学报,2007,16(1):47-51.
[12] 李亚,耿蕾,刘建秀.中国结缕草属植物抗盐性评价[J].草地学报,2004,12(1):8-11.
[13] 何秋,刘建秀,侯庆树.南京地区结缕草属植物锈病病原物鉴定及其发病规律的初步研究[J].江苏农业科学,2006,6:155-158.
[14] 王丹,宣继萍,郭海林,等.结缕草的抗寒性与体内碳水化合物、脯氨酸、可溶性蛋白季节动态变化的关系[J].草业学报,2011,20(4):98-107.
[15] 王舟,刘建秀.DREB/CBF类转达录因子研究进展及其在草坪草和牧草抗逆基因工程中的应用[J].草业学报,2011,20(1):222-236.
[16] 章元明,盖钧镒,王建康.利用B1和B2或F2群体鉴定数量性状主基因+多基因混合遗传模型并估计其遗传效应[J].生物数学学报,2000,15:358-366.
[17] 盖钧镒,章元明,王建康.植物数量性状遗传体系[M].北京:科学出版社,2003.
[18] 王建康,盖钧镒.利用杂种F2世代鉴定数量性状主基因-多基因混合遗传模型并估计其遗传参数[J].遗传学报,1997,24:432-440.
[19] 侯北伟,窦秉德,章元明,等.小麦雌性育性的主基因+多基因混合遗传分析[J].遗传,2006,28(12):1567-1572.
[20] 张立平,赵昌平,单福华,等.小麦光敏雄性不育系BS210育性的主基因+多基因混合遗传分析[J].作物学报,2007,33(9):1553-1557.
[21] 王庆钰,朱立宏,盖钧镒,等.水稻广亲和性遗传的主基因-多基因混合模型分析[J].遗传,2004,26(6):898-902.
[22] 罗庆云,於丙军,刘友良,等.栽培大豆耐盐性的主基因+多基因混合遗传分析[J].大豆科学,2004,23(4):239-243.
[23] 张洁夫,戚存扣,浦惠明,等.甘蓝型油菜花瓣缺失性状的主基因+多基因遗传分析[J].中国油料作物学报,2007,29(3):227-232.
[24] 郑轶琦,臧国长,郭海林,等.假俭草杂交后代生殖性状遗传及相关性分析[J].草业学报,2011,20(2):283-289.
