东大河林区金露梅种群空间分布格局沿海拔梯度的变化
2012-08-20刘建泉孙建忠
刘建泉,孙建忠
(1.甘肃祁连山国家级自然保护区管理局,甘肃 张掖734000;2.甘肃祁连山国家级自然保护区东大河保护站,甘肃 金昌737200)
种群分布格局的分析是研究种群特征、种内和种间关系以及种群与环境关系的重要手段,一直是生态学研究热点之一[1]。空间分布格局是植物种群生物学特性对环境生长适应和选择的结果,通常反映一定环境因子对个体行为、生长和生存的影响[2-3],是由种群特性、种群关系和环境条件的综合影响所决定的[4-5]。
金露梅(Potentilla fruticosa)灌丛在我国东北、华北、西北、西南各地均有分布;生长于海拔2 000m 以上的草地、草原及高山灌丛中[6],是高寒落叶灌丛的典型代表植物之一[7]。其广泛分布于青藏高原海拔2 300~4 500m的山地阴坡、半阴坡及洪积扇上[4,7]。东大河林区的金露梅灌丛多分布于亚高山灌丛带、森林草原带、灌丛草原带、山地荒漠草原带的阴坡、半阴坡、阳坡和半阳坡,发挥着重要的水源涵养、保持水土、保护生物多样性等多种生态功能。目前,有关金露梅灌丛的研究多集中在干扰生态学方面[8-12],而群落或种群生态学的研究鲜见报道[12-13]。本研究对东大河林区金露梅种群空间格局进行分析,旨在探讨种群的数量特征和与环境的关系,为金露梅灌丛的保护和恢复提供依据。
1 材料与方法
1.1 研究区自然概况 东大河林区位于祁连山中部的冷龙岭北麓,永昌县境内,地理位置101°24′~102°10′E,38°01′~38°10′N,属东大河、西大河中游,每年为镍都金昌市提供0.72亿m3的水源,是石羊河流域重要的水源涵养林区。据永昌县气象站资料,年平均气温1.9℃,最高温达31℃,最低温为-25℃,年降水量385.2mm,年蒸发量1 258.3mm,年均相对湿度50%~60%。土壤有山地灰褐土、山地栗钙土、亚高山草甸土等土壤类型;主要植被有分布于海拔2 500~3 400m的青海云杉(Picea crassifolia)林,分布于海拔2 500~3 600 m的金露梅灌丛,分布于海拔3 200~3 600m的鬼箭锦鸡儿(Caragana jubata)和柳(Salixspp.)灌丛,以及分布于沟谷地的小檗(Berberis spp.)、茶镳子(Ribes spp.)、栒子(Cotoneaster spp.)等组成的落叶阔叶灌丛。
1.2 样地调查 在多次踏查的基础上,2008年8月下旬选择东大河林区老人头资源管护站设置样带。老人头资源管护站所在山脉大致呈西南―东北走向,坡向大致呈东南方向,用GPS定位,沿海拔梯度设置样地5块,面积为8m×8m,每块样地内划分16个2m×2m的格子样方,采用植被调查方法,对每个样方植物的种类、多度、盖度[14]、高度等指标进行调查,同时记录样地自然环境(表1)。

表1 东大河林区金露梅灌丛样地概况Table 1 Brief introduction of Potentilla fruticosa shrub in Dongdahe forest
1.3 数据处理 测定植物种群分布格局的数学模型较多[4,15],其测定结果可能不完全相同,但综合分析各模型测定结果,一般可获得更可靠的结论[3]。分别按样地统计金露梅种群盖度,用格局指数C分析种群的分布格局[3,5,12,14-28],计算公式见参考文献[3,5-6,15-28]。分别用丛生指数(I)、Cassie指标(CA)、平均拥挤度(m*)、聚块性指标(L)、负二项参数(K)、扩散型指数(Is),共6个聚集强度指数分析金露梅种群的聚集强度,计算公式见参考文献[5,15-28]。当种群为聚集分布时,计算聚集均数:
式中,γ0.05为自由度等于2k时0.05水平下X2分布的内插和外插值。当λ<2时,造成聚集分布的原因是环境的作用引起;当λ≥2时,造成种群聚集分布的原因是种群生物学特性与环境的共同作用或其中的一个因素引起的[17-18,23]。
数据处理及分析采用 Micorsoft Excel 2007软件。
2 结果与分析
2.1 金露梅种群基本特征的变化 随着海拔升高,金露梅种群的平均高度、平均盖度和平均多度逐渐增加,在海拔3 017m时平均高度达到最大(29.75cm),随后下降;海拔2 990m 时平均盖度达到最大(5.94%),随后呈下降趋势;海拔2 842m时平均多度达到最大(3.25),在海拔3 140m 开始下降。种群在东南坡的适宜分布海拔高度大致为2 900~3 100m(图1)。
2.2 金露梅种群的空间分布格局分析 经过计算,金露梅种群格局指数 C=8.256 9,t=70.022 61>t0.05=2.645 7,呈聚集分布;聚集强度指数计算结果,丛生指数I=7.256 9、Cassie指标CA=1.878 8、平均拥挤度 m*=11.119 4、聚块性指标L=2.878 8、负二项参数 K=0.532 3,种群表现出很强的聚集特性。聚集均数=1.150 2<2.000 0,造成聚集分布的原因主要是金露梅种群的生存环境引起的。
2.3 金露梅种群的格局指数沿海拔高度的变化 不同海拔高度金露梅种群的空间分布格局都呈聚集分布格局(表2)。统计检验结果显示,除82703号样地不符合聚集分布外,其他样地均符合聚集分布,但82703号样地有明显的向聚集分布逼近的趋势;随着海拔升高,格局指数逐渐增加,在海拔2 990m处达到最大,随后出现下降和波动。聚集均数<2.000 0,造成聚集分布的原因主要是金露梅种群的生存环境引起的,但在海拔2 990m处的聚集均数为1.883 9,逼近2.000 0,金露梅种群的生物学特性也在逐渐影响种群的空间分布格局。
2.4 金露梅种群的聚集强度指数沿海拔高度的变化 不同海拔高度金露梅种群聚集强度指数的计算结果(表3)表明,随着海拔升高,负二项参数K逐渐减小,在海拔2 990m处达到最小,随后波动上升;Cassie指标CA、David和Morre指标逐渐增大,即随着海拔升高,种群聚集强度逐渐增大,在海拔2 990m处达到最大,随后波动下降,聚块性指数L则明显反映出种群扩散趋势和聚集趋势之间的波动,由2 990m向上和向下均表现为先呈扩散趋势,再呈聚集趋势。

图1 金露梅种群平均高度、平均盖度和平均多度随海拔高度的变化Fig.1 Change of average height,average coverage and average abundance of Potentilla fruticosa population along elevation
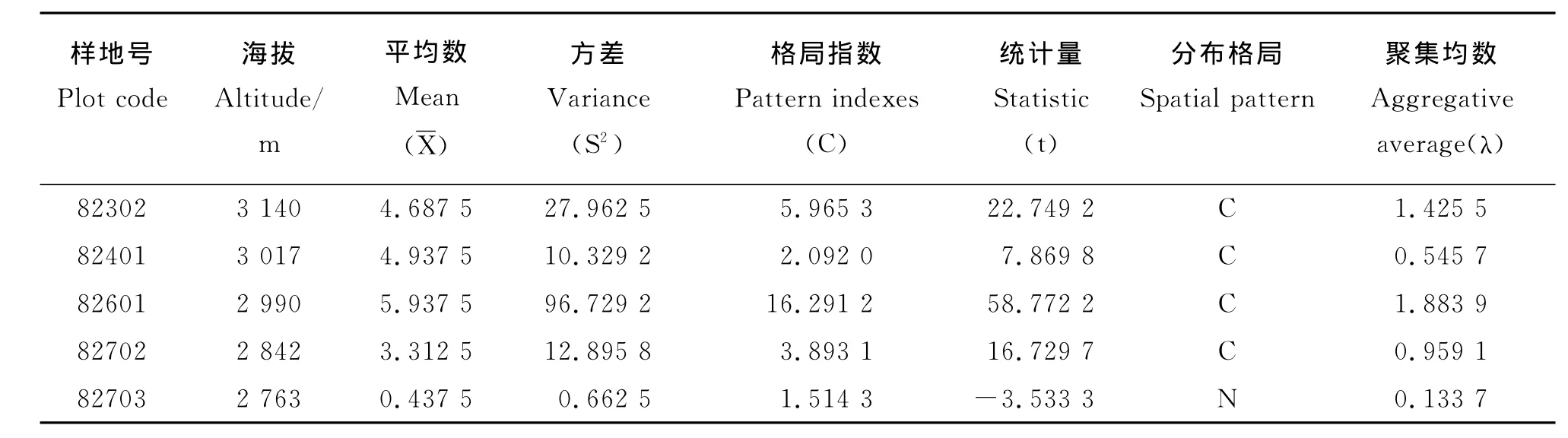
表2 东大河林区金露梅种群在不同海拔的格局指数Table 2 Pattern indexes of Potentilla fruticosa population at different elevation in Dongdahe forest

表3 东大河林区金露梅种群在不同海拔的聚集强度指数Table 3 Intensity indexes of Potentilla fruticosa population at different elevation in Dongdahe forest
3 讨论与结论
随着海拔升高,金露梅种群的平均多度、平均盖度、平均高度分别在海拔2 842、2 990、3 017m 处达到最大;金露梅种群在发育过程中,首先以增加多度来改善和维持群落环境,保证种群发展的需要;随着种群的发展,通过扩展空间、增加盖度满足对环境资源的需求,当环境资源不足以满足金露梅种群发展需要时,种群盖度降低,高度也随之下降,多度在维持一定的水平后也逐步降低。金露梅灌丛主要分布于海拔2 950~3 700m的山麓平缓起伏的阶地、潮湿河滩地、沟谷地、坡度<30°的山地阴坡和高海拔的山地阳坡[7],在东大河林区,金露梅种群在东南坡的适宜分布海拔高度为2 900~3 100m。
种群分布格局是物种与环境长期以来相互作用的结果,它不仅受物种本身的生物学特性和竞争排斥的影响,而且与物种的生境条件(包括土壤、地形、地貌)等 有 密 切 关 系[2,15,17]。 种 群 的 空 间 分 布 格 局在不同发育阶段、不同的生境条件下有明显差异[2,5,25]。金露梅种群总体上呈聚集分布,随着海拔升高,格局指数逐渐增加,在海拔2 990m处达到最大,随后出现下降和波动;聚集强度也逐渐增大,在海拔2 990m处达到最大,随后波动下降。造成聚集分布主要是金露梅种群的生存环境引起的,在东南坡、金露梅种群的适宜分布范围以内,种群的生物学特性对空间格局有一定的影响[17-19];适宜分布范围以外,主要是生存环境造成金露梅种群的聚集分布。聚集分布有利于金露梅种群发挥群体效应,形成适合种群自身生长的小环境,抵御不良环境和外界的干扰,从而维持种群的发展。
[1] 王本洋,余世孝.种群分布格局的多尺度分析[J].植物生态学报,2005,29(2):235-241.
[2] 张金屯,孟东平.芦芽山华北落叶松林不同龄级立木的点格局分析[J].生态学报,2004,24(1):35-40.
[3] 杨梅,林思祖,曹光球.人为干扰对常绿阔叶林主要种群分布格局的影响[J].中国生态农业学报,2007,15(1):9-11.
[4] 苏爱玲,徐广平,段吉闯,等.祁连山金露梅灌丛草甸群落结构及主要种群的点格局分析[J].西北植物学报,2010,30(6):1231-1239.
[5] 苏志尧,吴大荣,陈北光.粤北天然林优势种群结构与空间格 局 动 态 [J].应 用 生 态 学 报,2000,11(3):337-341.
[6] 郑健,郑勇奇,苑林,等.金露梅扦插繁殖技术研究[J].林业科学研究,2007,20(5):736-738.
[7] 于应文,胡自治,张德罡,等.金露梅灌丛净第一性生产力[J].草业学报,2000,9(4):33-39.
[8] 盛海彦,曹广民,李国荣,等.放牧干扰对祁连山高寒金露梅灌丛草甸群落的影响[J].生态环境学报,2009,18(1):235-241.
[9] 蒲小鹏,徐长林,刘晓静.放牧利用对金露梅灌丛土壤理化性质的影响[J].甘肃农业大学学报,2004,39(1):39-41.
[10] 刘发央,徐长林,龙瑞军.牦牛放牧强度对金露梅灌丛草地群落物种多样性的影响[J].草地学报,2008(6):613-618.
[11] 王爱东,尚占环,鱼小军,等.东祁连山北坡高寒灌丛草地围栏与放牧干扰下CO2释放速率的比较研究[J].甘肃农业大学学报,2010,45(1):120-124.
[12] 盛海彦,张春萍,曹广民,等.放牧对祁连山高寒金露梅灌丛草甸土壤环境的影响[J].生态环境学报,2009,18(3):1088-1093.
[13] 孙建忠,张燕,刘建泉,等.东大河林区金露梅灌丛种间关系分析[J].甘肃科技,2010,26(7):155-157.
[14] 温庆可,张增祥,刘斌,等.草地覆盖度测算方法研究进展[J].草业科学,2009,26(12):30-36.
[15] 茹文明,张桂萍,毕润成,等.濒危植物脱皮榆种群结构与分布格局研究[J].应用与环境生物学报,2007,13(1):14-17.
[16] 尹爱国,苏志尧,李彩虹.广东石门台自然保护区山顶矮林优势种群分布格局及动态[J].生态学杂志,2006,25(1):55-59.
[17] 刘建泉.祁连山保护区青海云杉种群分布格局的研究[J].西北林学院学报,2004,19(2):152-155.
[18] 郝虎,刘兴明,宋采福,等.祁连山北坡苔藓-青海云杉林优势种空间分布格局研究[J].甘肃林业科技,2005,30(4):9-12.
[19] 刘建泉,屈永才,王学福,等.青海云杉群落空间结构特征 研 究 [J].干 旱区 资 源 与 环 境,2009,23(7):121-125.
[20] 刘足根,朱教君,袁小兰,等.辽东山区次生林主要树种种群结构和格局[J].北京林业大学学报,2007,29(1):12-18.
[21] 张金屯.数量生态学[M].北京:科学出版社,2004:245-252.
[22] 江洪.云杉种群生态学[M].北京:中国林业出版社,1992:8-26.
[23] 丁岩钦.昆虫数学生态学[M].北京:科学出版社,1994:44-50.
[24] 王得祥,刘建军,李登武,等.秦岭山地华山松林群落学特 征 研 究 [J].应 用 生 态 学 报,2000,15(3):357-362.
[25] 吴大荣,苏志尧,李秉滔,等.福建三明莘口青钩栲种群结构和空间分布格局动态初步研究[J].林业科学,2000,36(3):27-32.
[26] 王海为,游水生,王小明.锥栗林主要种群空间分布格局[J].福建林学院学报,2004,24(3):258-261.
[27] 李毅,胡自治,王志泰.东祁连山高寒地区山生柳种群分布格局研究[J].草业学报,2002,11(3):48-54.
[28] 兰国玉,雷瑞德.植物种群空间分布格局研究方法概述[J].西北林学院学报,2003,18(2):17-21.
