钙对茉莉酸甲酯诱导番茄抗灰霉病及防御酶活性的调控作用
2012-08-08余朝阁李天来张亢亢刘志恒李琳琳
余朝阁 李天来 张亢亢 刘志恒 李琳琳 周 娣
(1沈阳农业大学园艺学院,设施园艺省部共建教育部重点实验室,辽宁沈阳 110161;2沈阳农业大学植物保护学院,辽宁沈阳 110161;3沈阳农业大学图书馆,辽宁沈阳 110161)
茉莉酸(Jasmonic Acid,JA)及其甲酯(Methyl Jasmonate,MeJA)是广泛存在于植物体中的重要内源信号物质,它不仅参与植物抵抗生物胁迫或非生物胁迫的信号传递过程,也是重要的外源激发子,可诱导植物对多种逆境胁迫发生抗性反应(Turner et al.,2002;Thaler et al.,2004)。大量研究发现,植物受机械损伤、病菌侵染或昆虫取食等逆境胁迫后,JA类物质含量大幅升高;外源JA或MeJA则可诱导多种植物抗逆性增强;而茉莉酸缺失或不敏感突变体的抗逆性往往显著低于其相应的野生株(Kazan & Manners,2008;Browse,2009)。
钙作为植物必需营养元素和重要的胞内第二信使,参与植物对多种逆境抗性信号传递途径的调控(Lecourieux et al.,2002;Dodd et al.,2010)。JA可诱导蚕豆叶片细胞质中钙离子浓度〔Ca2+〕cyt的迅速上升,从而诱导气孔关闭,而Ca2+螯合剂EGTA和质膜Ca2+通道的抑制剂硝苯吡啶(Nifedipine,NIF)则阻断或减弱诱导的气孔关闭效应(刘新 等,2005);另外,JA也诱导拟南芥细胞的钙动员,这一过程既包括胞外Ca2+的流入,也包括胞内钙库中Ca2+的释放(孙清鹏 等,2010);但也有研究认为,JA对番茄叶肉细胞中〔Ca2+〕cyt没有影响(Moyen et al.,1998)。
对比不同研究结果发现,钙在JA信号传递途径中的作用还存在争议,机制尚不明确。本试验系统探索钙对MeJA诱导番茄抗灰霉病和几种重要防御酶活性的影响,为揭示JA诱导番茄抗病性与Ca2+的关系提供依据。
1 材料与方法
1.1 供试材料
番茄(Lycopersicon esculentumMill.)品种为L402,灰霉菌(Botrytis cinerea)由田间分离所得。供试化学物质MeJA购自Sigma公司,CaCl2、LaCl3和EGTA均购自国药集团化学试剂有限公司。
1.2 材料培养与处理
番茄采用穴盘育苗,三叶期定植于直径为12 cm的塑料钵中。六叶期将幼苗分为5组,其中4组用3 mmol·L-1的MeJA(先用乙醇助溶,然后溶解于蒸馏水,终浓度MeJA为3 mmol·L-1,乙醇体积比为0.1%)涂抹第3片叶至完全润湿,对照组则涂抹0.1%的乙醇;晾干后(约1.5 h),对照和其中1组MeJA处理喷施蒸馏水至完全润湿(CK和MeJA),其余3组分别喷施等量的20 mmol·L-1的 CaCl2(MeJA+Ca)、3 mmol·L-1的 LaCl3(MeJA+La)和 5 mmol·L-1的 EGTA(MeJA+EGTA)。5 d后用每毫升含1×106个灰霉菌孢子的悬液接种各处理植株的第3片叶前5裂叶,接种采用微量注射法(Audenaert et al.,2002)。每处理5株,3次重复。
1.3 测定指标及方法
番茄幼苗接种后第5天调查发病程度并计算病情指数,病害分级标准参照李洪连和徐敬友(2001)的方法。诱导处理后0、1、2、3、5 d分别取第3片叶,在液氮速冻后于-86 ℃下保存,用于植物防御酶活性测定。苯丙氨酸解氨酶(PAL)、多酚氧化酶(PPO)、过氧化物酶(POD)、过氧化氢酶(CAT)和超氧化物歧化酶(SOD)的提取和活性测定参照余朝阁(2007)的方法。
1.4 数据处理
试验数据均用SPSS软件进行方差分析和多重比较。
2 结果与分析
2.1 钙对MeJA诱导番茄叶片抗灰霉病的影响
MeJA显著提高番茄抗灰霉病程度,MeJA单独处理后,番茄叶片病情指数比对照低32.1%。外源钙离子进一步增强MeJA诱导的抗病性,MeJA与CaCl2共同处理(MeJA+Ca)的病情指数比MeJA单独处理的病情指数降低15.1%,达到显著水平。而MeJA的诱导抗病性作用可被质膜钙通道抑制剂LaCl3和Ca2+螯合剂EGTA完全抑制,MeJA+EGTA和MeJA+La处理的病情指数与对照均无显著差异,但EGTA的抑制作用显著强于 LaCl3(图 1)。上述结果表明,MeJA可诱导番茄抗灰霉病,外源钙离子增强MeJA诱导的抗病性,缺钙处理则抑制MeJA诱导的抗病性,钙可能参与MeJA诱导番茄抗灰霉病的调控过程。

图2 钙对MeJA诱导番茄叶片中PAL活性的影响
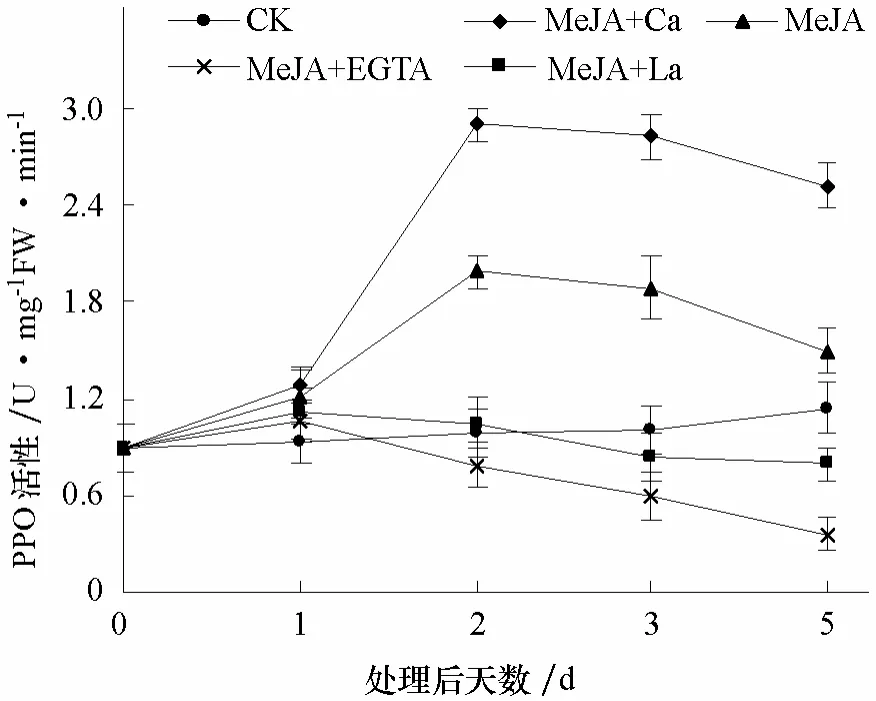
图3 钙对MeJA诱导番茄叶片中PPO活性的影响
2.2 钙对 MeJA诱导番茄叶片中防御酶活性的影响
2.2.1 PAL活性 各处理番茄叶片中 PAL活性在处理后第1天升高,并均高于对照。其中,MeJA单独处理的PAL活性第2天迅速上升并达到高峰,然后缓慢下降,但均显著高于对照。MeJA+Ca处理的PAL活性也在第2天迅速上升,且第2~5天始终保持较高水平,并显著高于MeJA单独处理。MeJA+EGTA处理的PAL活性在第1天升高后,第1~3天无显著变化,至第5天则降至与对照相当的水平;MeJA+La处理的PAL活性变化趋势和MeJA单独处理类似,但第2~5天PAL活性略低于MeJA单独处理(图 2)。上述结果说明,外源钙离子促进MeJA对番茄 PAL活性的诱导,而 EGTA和LaCl3则抑制 MeJA诱导番茄 PAL活性,且EGTA的抑制作用强于LaCl3。
2.2.2 PPO活性 各处理番茄叶片中 PPO活性在处理后第1天均无显著变化。其中,MeJA单独处理后,PPO活性第2天迅速升高并达到高峰,然后缓慢下降,到第5天降至对照水平。MeJA+Ca处理的PPO活性与MeJA单独处理的变化趋势类似,但在第 2~5天始终显著高于MeJA单独处理。MeJA+EGTA处理的番茄叶片中 PPO活性从第 2天开始略低于对照,之后继续下降,第5天时显著低于对照;MeJA+La处理后3 d内,PPO活性与对照无显著差异,第5天显著低于对照(图3)。说明外源钙离子促进MeJA对番茄PPO活性的诱导,而EGTA和LaCl3则抑制MeJA对番茄PPO活性的诱导作用,且EGTA的抑制程度强于LaCl3。
2.2.3 POD活性 各处理番茄叶片中 POD活性在处理后第1天均无显著变化,第2天迅速升高,第3天继续升高,且在第2~3天内均显著高于对照。其中,MeJA+Ca和MeJA单独处理的 POD活性变化趋势整体上一致,处理后第5天仍然继续快速升高;但是,在第2~5天内,MeJA+Ca处理的 POD活性均显著高于 MeJA单独处理。MeJA+La处理的POD活性第5天也继续升高,但显著低于MeJA单独处理;MeJA+EGTA处理第3~5天无显著变化,且第5天和对照无显著差异(图4)。上述结果说明,外源钙离子也促进 MeJA对番茄 POD活性的诱导作用,而缺钙处理对MeJA诱导番茄POD活性具有一定的抑制作用。
2.2.4 CAT活性 MeJA单独处理后3 d内,CAT活性与对照无显著差异,第 5天有所升高,并显著高于对照。MeJA+Ca处理后1~5 d内,CAT活性始终保持缓慢上升,至第5天显著高于对照和其他各处理。MeJA+EGTA和MeJA+La两个处理的CAT活性均在第1天迅速升高;MeJA+EGTA处理的CAT活性在第2天和第1天之间无显著变化,第2~5天缓慢下降;而MeJA+La处理的CAT活性在第2天迅速下降至对照水平以下,第2~5天缓慢上升至对照水平(图 5)。整体看来,外源钙离子对MeJA诱导番茄CAT活性的影响无明显的规律性。
2.2.5 SOD活性 各处理 SOD活性均呈先升高、后降低、再升高的趋势。但 MeJA+Ca和MeJA+EGTA处理第1~3天缓慢下降至对照水平以下,第3~5天升高至对照水平以上。而MeJA+La和 MeJA单独处理第 1~2天迅速下降至对照水平以下,第2~5天缓慢升高至对照水平(图6)。上述结果显示,外源钙离子对MeJA诱导番茄SOD活性的作用也不确定。

图4 钙对MeJA诱导番茄叶片中POD活性的影响
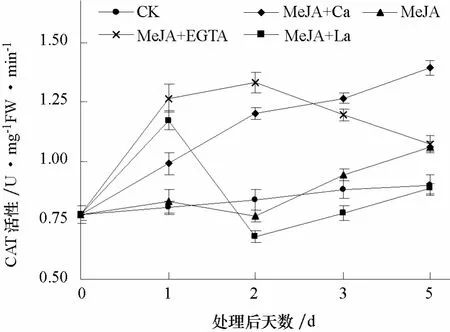
图5 钙对MeJA诱导番茄叶片中CAT活性的影响
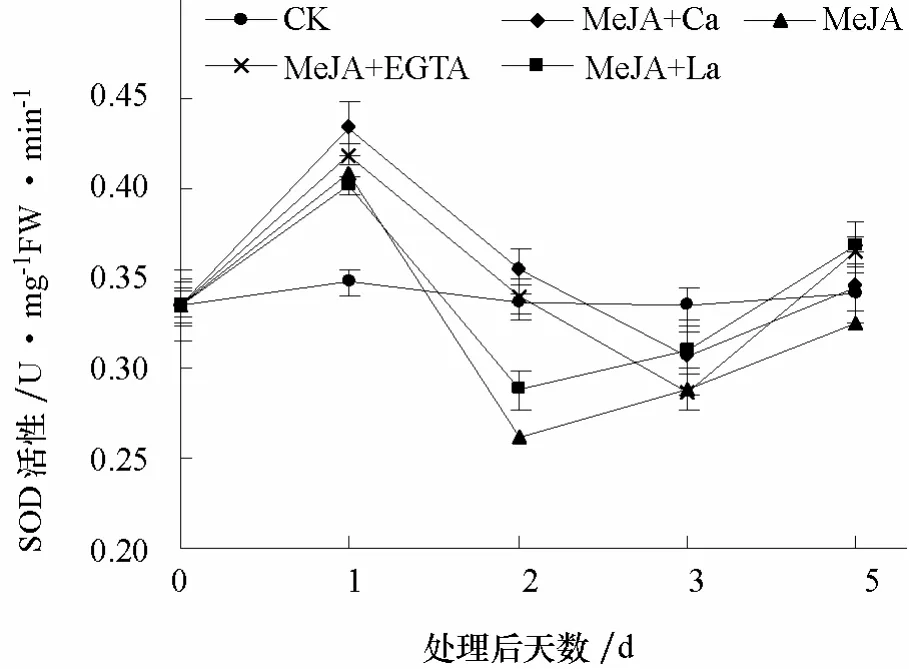
图6 钙对MeJA诱导番茄叶片中SOD活性的影响
3 结论与讨论
本试验结果表明,外源MeJA可诱导番茄抗灰霉病能力增强,也诱导番茄叶片中PAL、PPO和POD等防御酶活性提高。外源钙离子进一步增强MeJA诱导的抗病性及上述防御酶活性;钙螯合剂EGTA或质膜钙通道抑制剂LaCl3均抑制MeJA诱导番茄的抗病性和上述防御酶活性。而钙对MeJA诱导番茄叶片中CAT和SOD活性的影响未发现明显的规律性。所以认为,钙对MeJA诱导番茄抗灰霉病的调控可能主要与其对MeJA诱导番茄叶片中PAL、PPO和POD活性的影响有关。
尽管钙和茉莉酸类物质在植物抵抗逆境胁迫中的作用已有大量系统深入研究,也有部分研究发现 Ca2+参与JA介导的信号转导过程;但是,对Ca2+和JA在植物抗逆性中的相互关系的研究并不多见,特别是钙对JA类物质诱导的植物抗病性和诱导植物防御酶反应的影响研究,更是鲜见报道。本试验探明了外源钙离子在 MeJA诱导番茄抗灰霉病中的作用,并系统探索了钙对MeJA诱导番茄5种重要防御酶活性的影响,初步明确外源钙离子增强MeJA诱导番茄抗灰霉病程度,与其对MeJA诱导番茄PAL、PPO和POD3种主要防御酶活性的调控有关。该试验结果将为揭示钙对MeJA诱导番茄抗病性的调控机制研究奠定基础。
PAL、PPO和POD是催化植物体中酚类物质和木质素形成的关键酶,而酚类物质和木质素则是植物体中重要的抗病物质;钙参与调节 MeJA诱导上述防御酶活性,推测钙也可能进一步影响MeJA诱导番茄酚类物质和木质素等抗病物质的积累。这也将是进一步研究的重要内容。
钙对MeJA诱导番茄叶片中CAT和SOD活性未发现规律性影响。CAT和SOD是重要的抗氧化酶,可以清除植物体内活性氧;而活性氧一方面可以激活植物防御系统,另一方面,其大量积累也将对植物自身产生伤害作用。钙对MeJA诱导番茄CAT和SOD活性影响的不确定性,可能与植物体中活性氧作用的两面性和复杂性有关。
李洪连,徐敬友.2001.农业植物病理学实验实习指导.北京:中国农业出版社:150.
刘新,石武良,张蜀秋,娄成后.2005.Ca2+参与茉莉酸诱导蚕豆气孔关闭的信号转导.实验生物学报,38(4):297-301.
孙清鹏,于涌鲲,万善霞,赵福宽,郝玉兰.2010.胞外及胞内Ca2+共同参与拟南芥中茉莉酸诱导的钙动员.中国农业科学,43(5):942-948.余朝阁.2007.钙对化学诱抗剂诱导番茄抗灰霉病及其相关防卫反应的调控作用〔博士论文〕.沈阳:沈阳农业大学:36-38.
Audenaert K,De Meyer G B,Hofte M M.2002.Abscisic acid determines basal susceptibility of tomato toBotrytis cinereaand suppresses salicylic acid-dependent signaling mechanisms.Plant Physiology,128(2):491-501.
Browse J.2009.Jasmonate passes muster:a receptor and targets for the defense hormone.Annu Rev Plant Biol,60:183-205.
Dodd A N,Kudla J,Sanders D.2010.The language of calcium signaling.Annu Rev Plant Biol,61:593-620.
Lecourieux D,Mazars C,Pauly N,Ranjeva R,Pugin A.2002.Analysis and effects of cytosolic free calcium increases in response to elicitors inNicotiana plumbaginifoliacells.The Plant Cell,14:2627-2641.
Moyen C,Hammond-Kosack K E,Jones J,Knight M R,Johannes E.1998.Systemin triggers an increase of cytoplasmic calcium in tomato mesophyll cells:Ca2+mobilization from intra-and extracellular compartments.Plant,Cell & Environmet,21:1101-1111.
Kazan K,Manners J M.2008.Jasmonate signaling:toward an integrated view.Plant Physiology,146:1459-1468.
Thaler J S,Owen B,Higgins V J.2004.The role of the jasmonate response in plant susceptibility to diverse pathogens with a range of lifestyles.Plant Physiol,135:530-538.
Turner J G,Ellis C,Devoto A.2002.The jasmonate signal pathway.The Plant Cell,Supplement:153-164.
