基于全极化Radarsat-2数据的水稻生物量估算模型
2012-07-10张晓倩刘湘南
张晓倩,刘湘南,谭 正
(中国地质大学(北京)信息工程学院,北京100083)
水稻是人类主要的粮食作物,而我国作为人口大国,水稻种植面积约占全世界五分之一,水稻总产量居于世界首位。如何即时获取大面积水稻生长信息,从而指导水稻生产成为国家决策的重要依据,也是亟待解决的科学问题。对于大面积的水稻监测,传统的依靠地面监测显然不能满足需求,卫星遥感监测成为水稻监测的重要手段。然而,由于我国水稻种植区域多分布在云雨频繁的热带、亚热带地区,光学遥感卫星无法获取实时的高清晰水稻遥感影像[1],对于植被垂直方向上的信息反应不足,而雷达遥感采取侧视成像,工作在微波波段(1mm~1m),波长高于光学遥感的雷达波可穿云透雾[2],不仅受气候影响较少,对植被等地物也有一定的穿透能力,利用雷达遥感进行生物学参数反演显现出一定优势。Radarsat-2为2007年12月14日发射的雷达成像卫星,具有3m高分辨率成像能力,全极化(HH、VH、HV、VV)信息将进一步提高水稻生物参数反演精度,近几年该数据用于水稻研究鲜有应用。
大量研究表明,雷达后向散射系数对水稻生物量等生物参数比较敏感,国内外已就雷达电磁波与水稻间的作用机理发展很多水稻后向散射模型,以反演水稻生物学参数。Le Toan等[3,4]采用多时相单极化SAR数据分析水稻后向散射特征进行水稻识别和水稻参数反演;Inoue等[5]采用水稻生长期内多频、全极化和不同入射角数据研究其后向散射信号与水稻冠层生物参数的关系;邵芸等[6,7]利用多时相、多模式的雷达数据,分析了水稻的时域散射特性与水稻年龄的关系,提出可进行水稻识别估产的最佳时像以及进行水稻监测的最优雷达参数;董彦芳等[8,9]利用多时相ASAR数据进行水稻长势监测,改进Sun和Simonett提出的连续冠层模型,实现了对水稻后向散射的模拟,尝试水稻生物参数反演。
基于雷达遥感能够穿云透雾,更易获取植被立体信息的优势,本文旨在建立水稻生物量与雷达数据关系。选取具有一定物理意义的半经验水云模型、回归模型,分析Radarsat-2数据四种极化方式时域变化特征,对不同极化及其组合分别建模,通过对比寻找可进行水稻生物量反演的最优极化方式与模型。
1 试验数据采集和处理
1.1 试验区域数据采集
研究区域位于江苏省苏州市东桥镇(31°25′N,120°31′E)。苏州是传统的水稻种植区,水稻种植面积不少于100万亩,该区为亚热带湿润季风气候区,水稻种植季节云雨天气较多。东桥镇试验区域设置2个样区,大小均为1 hm2~1.5hm2,其中1个样区位于废弃化工厂旁,重金属污染导致水稻生长状况低于健康水稻,另一样区为健康水稻。于2009年7月2日、7月28日、8月28日、9月28日分别进行以采集水稻样本获取生物参数的野外遥感调查,采用拔节到抽穗期即7月28日、8月28日数据进行研究。每个样区选取10个1 m×1m大小样点,每样点采集地表活株样本称重即可得到水稻鲜生物量,通过GPS记录样本点的经纬度,便于将实测数据与雷达影像一一对应。
1.2 雷达数据获取和预处理
卫星数据为2007年12月14日发射的商用雷达卫星Radarsat-2获得的全极化雷达数据,传感器工作在C波段,雷达卫星数据的成像参数见表1,该影像分辨率为12m*8m,包含HH、HV、VH、VV四种不同极化方式数据。

表1 Radarsat-2数据成像参数
在雷达数据进行分析前,需要对其进行预处理。使用欧空局的NEST软件对影像进行辐射定标,使雷达影像DN值转换为后向散射系数(dB)便于定量使用。对影像进行多视操作以抑制原始图像上存在的固有噪声斑点,再采用3*3窗口的Lee滤波来对雷达四种极化影像进行去噪,处理后地物内部平滑,地物边缘锐化。依据地面实测数据经纬度在影像上选取控制点得到不同极化后向散射系数。
2 研究方法
2.1 水云模型描述
许多研究表明,雷达后向散射对水稻生物参数十分敏感[3-10]。可建立多极化雷达后向散射系数与水稻生物参数模型,便可利用雷达影像数据直接反演水稻生物参数。雷达后向散射受到诸多变量影响,如植被、土壤、地形以及雷达传感器本身。整个冠层的后向散射系数0可用基于辐射传输方程一阶解的半经验植被后向散射模型水云模型[5,11]来描述,该模型假定植被冠层由分布均匀且大小相等的微小水滴组成,认为后向散射系数分解为两部分:(1)冠层自身的体散射;(2)经过冠层衰减后土壤的散射。该模型表述为:
影像中后向散射系数以dB表征,故水稻后向散射系数(dB)也可描述如下:
对方程(4)求其反向解,则水稻生物量FW为:
其中,σ0为雷达影像不同极化后向散射系数,参数a、b、c可通过雷达影像后向散射系数与实测水稻生物量进行拟合得到。
2.2 二次多项式与指数模型描述
雷达后向散射系数与水稻生物参数有复杂的相关关系,简单的线性模型不能准确描述其关系[15],不宜选取,因此本文将建立雷达影像后向散射系数与水稻生物量估算经验模型,分别为二次多项式模型(8)和指数模型(9),与水云模型进行比较。FW为水稻生物量,σ0为雷达不同极化后向散射系数,公式中参数a、b、c分别由实测生物量与后向散射系数拟合得到。
3 结果与分析
3.1 雷达不同极化后向散射系数时域变化特征
本文选取水稻生长期内生物量累积较快的时期进行研究。2009年7月26日(下文均简化为7-26)为水稻拔节期,2009年8月26日(下文均简化为8-26)为水稻抽穗期,此时期为水稻生物量积累较快的时期。两景影像分别选取水稻区域30个控制点比较四种极化HH、HV、VH、VV后向散射系数变化情况(图1)。7-26水稻HH极化后向散射系数平均值为-10.219dB,VV极化后向散射系数平均值为-14.821dB;8-26水稻平均值为-7.276 dB,平均值为-9.506 dB,据图1中控制点的极化波动也可得到,对于同一地物,均有,而随着水稻植株的生长都表现出增加的趋势,两个时期比较平均增加3 dB增加5 dB。
两时相中交叉极化值均小于同极化值。理论上,对于同一地物微波后向散射其交叉极化HV、VH是相等的,HV、VH在不同波段跟不同入射角相关系数都极高[5],这在图1中也可得到验证。通过计算得出交叉极化HV与VH后向散射系数差值很小,7-26水稻HV极化后向散射系数平均值为-20.036dB,VH极化后向散射系数平均值为-19.893dB,相差0.143 dB;8-26水稻平均值为-16.352dB,平均值为-16.174dB,二者差值为0.178 dB。因而,研究中每个采样点都以平均数作为水稻交叉极化数据进行研究。
HH极化与VV极化比值HH/VV已证明在雷达影像分类中有较高的精度,同时对水稻生物量等参数变化较敏感[15]。研究中分别比较了归一化HH、VV、CROSS、HH/VV后向散射系数的标准差[13](表2),两个时相HH/VV标准差分别小于同时相其他三种极化标准差,表明HH/VV较其余极化更为稳定。基于以上考虑,研究中将HH/VV作为一种极化组合方式,与其他三种极化共同建模比较。
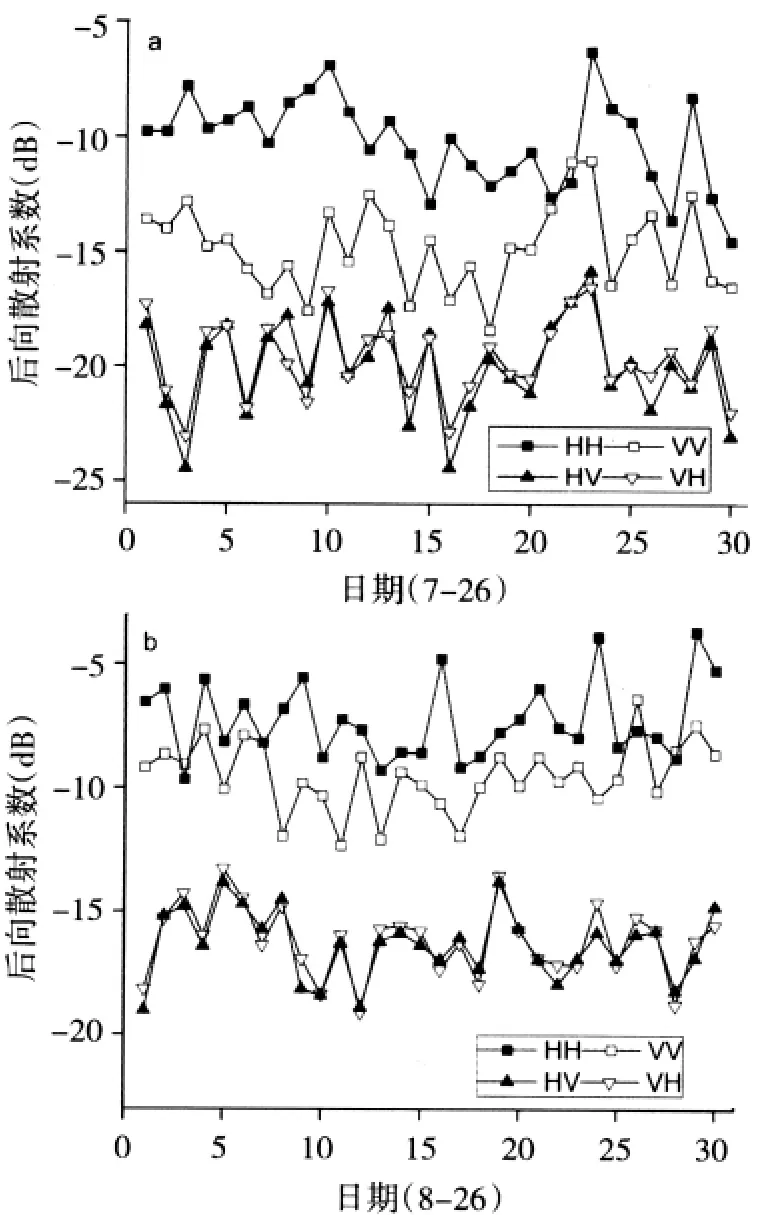
图1 水稻后向散射系数时域变化特征(a)7月26日和(b)8月26日
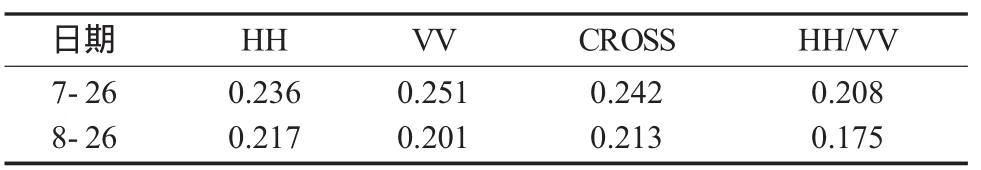
表2 不同时相归一化后向散射系数标准差
3.2 水云模型建模分析
将Radarsat-2影像后向散射系数与实测水稻生物量依据水云模型 FW=a*b-b*In(c-σ0)进行拟合,以确定参数 a、b、c的值。通过上节对雷达不同极化后向散射系数时域变化特征分析,在此将HH、VV、CROSS、HH/VV四种极化组合方式的后向散射系数(m2/m2)引入水云模型进行拟合比较。表3展示了HH、VV、CROSS、HH/VV四种极化方式建模情况。

表3 不同极化水云模型建模
HH极化跟CROSS极化情况下的拟合相关系数跟均方根误差相似,拟合相关系数分别为0.910、0.902,都具有较高的拟合精度;两种极化均方根误差分别为0.190、0.216,误差相对较小。这两种极化情形下,随着水稻生物量的积累,后向散射系数单调递增且增长迅速,后向散射系数越大生物量的积累越迅速,说明这两种极化对水稻生物量比较敏感。HH/VV极化情况下相关系数为0.893,拟合精度略低于上述两种极化,均方根误差0.287,明显高于上述两种极化,误差增加。生物量在HH/VV较小时增加缓慢,当HH/VV>5.5时,比值不再随着生物量的增加而增加,比值达到饱和,说明在水稻抽穗后比值对生物量不敏感,不宜采用水云模型模拟。VV极化情况下,拟合相关系数为0.748,拟合相关系数较低,均方根误差为0.435,与其余三种极化方式相比误差较大,随着VV极化后向散射系数变化对生物量敏感度降低,反演效果逊于 HH、CROSS、HH/VV。
3.3 二次多项式与指数模型建模分析
雷达后向散射系数与水稻生物参数有复杂的相关关系,简单的线性关系模拟精度较低不宜选取,因此本文选取常用的非线性回归模型二次多项式模型与指数模型对四种极化组合方式HH、VV、CROSS、HH/VV进行拟合比较。不同极化雷达后向散射系数与实测水稻生物量拟合为方程(8)、(9)的形式,见表4。
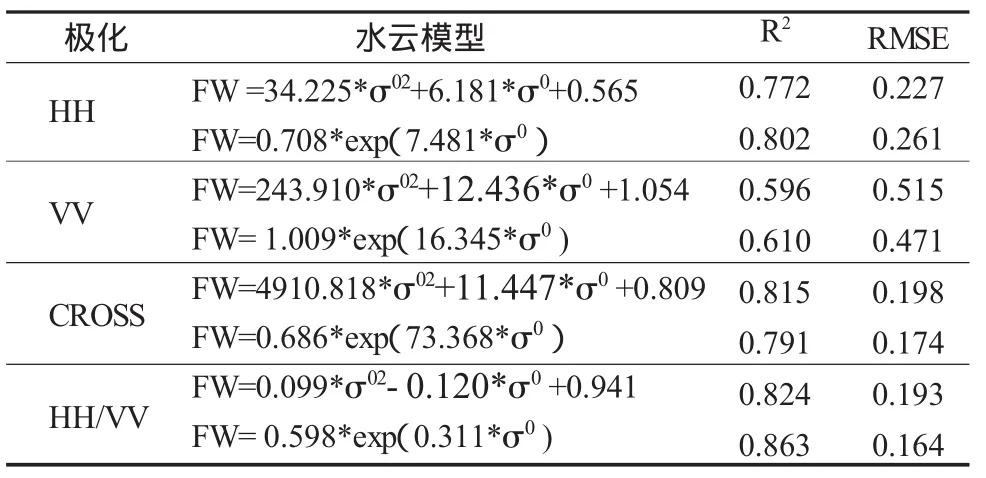
表4 回归模型比较
四种极化方式水云模型与非线性回归模型有相似的模拟结果。HH极化、CROSS极化非线性回归模型拟合相关系数均为0.9左右,但HH极化均方根误差较CROSS极化大。HH/VV两种回归模型相关系数最高,均方根误差相对较小。VV极化两种回归模型相关系数低,均方根误差高,模拟效果较其余三种极化都低。二次多项式跟指数模型相比,除交叉极化外,其余三种极化指数模型模拟效果较好,相关系数较高,均方根误差也较小。非线性回归模型与水云模型相比,HH极化、CROSS极化水云模型优于非线性回归模型,HH/VV则是非线性回归模型较好,VV极化三种模型模拟效果均最差。
3.4 精度分析
选取精度相对较高的水稻生物量反演模型,HH极化水云反演模型和HH/VV指数反演模型,分别比较水稻模拟生物量与实测生物量,分析模型模拟不同生物量水平的精度。研究表明,HH水云模型和HH/VV指数模型都表现出对于1.75 kg/m2~2.5kg/m2水平生物量模拟效果较好,RMSE分别为0.140kg/m2、0.121kg/m2。HH水云模型对低于 1.75kg/m2水平生物量模拟效果差,RMSE分别为0.218kg/m2,HH/VV指数模型对高于2.5kg/m2水平生物量模拟效果差,RMSE分别为0.175kg/m2。HH水云模型对低水平生物量模拟效果较差,这主要是由于在低水平生物量情况下,水稻密度较低,电磁波极易穿透水稻射入地表,此时后向散射系数值很大一部分来自于地表反射,由此产生了比水稻植株本身更强的回波信号。HH/VV指数模型对于高水平水稻生物量模拟效果差,主要是由于同极化比值随着生物量增加会趋近饱和,随着生物量增大,同极化比值增长缓慢,同样在HH/VV水云模型中HH/VV>5.5时比值的变化不能很好的反应生物量的增加。
通过HH水云模型和HH/VV指数模型计算大面积水稻生物量并尝试绘制水稻生物量结果分布图。裁剪2009年8月26日影像研究区附近大面积水稻区域,水稻外地物赋值白色,水稻生物量分布情况见图2。计算两分布图中水稻生物量为2.0 kg/m2-3.8kg/m2范围的像元个数分别占水稻像元总数的65.26%、53.94%。对于大面积的水稻生物量分布,受到影像水稻识别精度、不同区块水稻耕作方式等因素影响,精度有待于进一步验证。

图2 根据雷达影像得到水稻生物量分布图(a)HH水云模型和(b)HH/VV指数模型
4 结论和讨论
本研究比较了雷达C波段HH、VV、CROSS、HH/VV四种极化及组合方式的水云模型、二次多项式模型和指数模型反演水稻拔节到抽穗期的水稻生物量,总体取得比较好的效果。改进水云模型作为一种半经验模型,不仅具备一定的物理含义同时参数少易于反演,相对于纯理论模型与经验模型有一定优势。从反演结果来看,HH、CROSS、HH/VV水云模型都有不错的反演效果,而HH水云模型反演生物量尤佳,指数模型普遍优于二次多项式模型,HH/VV指数模型效果出众。将HH水云模型、HH/VV指数模型分别应用于影像大面积水稻田,为大面积的水稻监测提供可能。然而,一方面由于两个水稻样区受污染胁迫不同导致水稻生长差异,给本文分析结果的可靠性和普遍性造成一定影响;另一方面用于建模的两景影像入射角差异对结果也造成一定影响,因此本文结果还需更多的数据来验证与完善。
水稻生物量与LAI、稻龄、株高等参数有极高的相关关系[16,17],今后研究中可考虑在此基础上进行水稻LAI、稻龄、株高等参数反演研究。另外,多极化、多模式、特殊波段越来越突出了星载雷达的优势,同时也拓宽了雷达的应用范围[2],各种新模式数据的应用成为趋势。研究中尝试使用Radarsat-2多极化数据进行水稻生物参数反演,对于不同极化组合数据反演水稻生物参数提供可能。
[1]XiaoX M,Boles S,Liu J Y,et al.Mapping paddy rice agriculture in Southern China using multi-temporal MODIS images[J].Remote SensingofEnvironment,2005(95):480-492.
[2]唐鹏钦,姚艳敏,魏娜.合成孔径雷达水稻识别和监测研究进展[J].中国农学通报,2009,25(14):291-295.
[3]Le ToanT,LaurH,MouginE,et al.Multitemporal and dual-polarization observation of agricultural vegetation covers by X-band SAR images[J].IEEE Transactions of Geo-science and Remote Sensing,1989,27(6):709-718.
[4]Le Toan T,Ribbes F,Wang L-F,et al.Rice crop mapping and monitoring using ERS-1 data based on experiment and modeling results[J].IEEE Trans Geosci Remote Sensing,1997,35(1):41-56.
[5]Inoue Y,Kurosu T,Maeno H,et al.Season-long daily measurements of multifrequency (Ka,Ku,X,C,and L)and full-polarization backscatter signatures over paddy rice field and their relationship with biological variables[J].Remote Sensing of Environment,2002(81):194-204.
[6]Yun S,Fan X T,Liu H,et al.Rice monitoring and production estimation using multitemporal RADARSAT[J].Remote Sensing of Environment,2001(76):310-325.
[7]邵芸,廖静娟,范湘涛,等.水稻时域后向散射特性分析:雷达卫星观测与模型模拟结果对比[J].遥感学报,2002,6(6):440-450.
[8]董彦芳,孙国清,庞勇.基于ENVISATASAR数据的水稻监测[J].中国科学D辑地球科学,2005,35(7):682-689.
[9]董彦芳,庞勇,孙国清.ENVISAT ASAR数据用于水稻监测和参数反演[J].武汉大学学报(信息科学版),2006,31(2):124-127.
[10]申双和,杨沈斌,李秉柏,等.基于ENVISAT ASAR数据的水稻时域后向散射特征分析[J].农业工程学报,2009,25(增刊2):130-136.
[11]Attema E P W,Ulaby F T.Vegetation modeled as a water cloud[J].RadioScience,1978,(13)2:357-364.
[12]Kurvonen L,Pulliainen J,Hallikainen M.Retrieval of biomass in boreal forests from multitemporal ERS-1 and JERS-1 SAR images[J].IEEE Transactions on Geoscience and Remote Sensing,1999,37(1):198-205.
[13]何维.基于ASAR和生长模拟模型的水稻长势监测研究[D].北京:中国林业科学研究院,2007.
[14]黎夏,叶嘉安,王树功,等.红树林湿地植被生物量的雷达遥感估算[J].遥感学报,2006,10(3):387-396.
[15]De Jong SM S,Pebesma E J,Lacaze B.Above-ground biom assassessment of mediterranean forests using airborne imaging spectrometry:the DAIS peyne experiment[J].International Journal of Remote Sensing,2003,24(7):1505-1520.
[16]ChakrabortyM,ManjunathKR,PanigrahyS,etal.Ricecropparameter retrieval using multi-temporal,multi-incidence angle Radarsat SAR data[J].ISPRS Journal of Photogrammetry&Remote Sensing,2005,(59):310-322.
[17]Shen S H,Yang S B,Li B B,et al.A scheme for regional rice yield estimation using ENVISAT ASAR data[J].Sci China Ser D-Earth Sci,2009,52(8):1183-1194.
