干旱和复水对冬小麦光合产物分配格局的影响
2012-07-07谷艳芳丁圣彦高志英
谷艳芳,丁圣彦,高志英,邢 倩
(1.河南大学生态科学和技术研究所,河南开封 475001;2.河南大学生命科学学院,河南开封 475001)
1 前言
作物对水分胁迫及胁迫后复水的响应是一个十分复杂的问题,冬小麦生长发育过程中不同生育期由于其生长中心不同,常常对水分的需求和对胁迫的敏感性也不同[1,2]。黄淮海地区冬小麦在关键生育期,即拔节-灌浆期常出现降雨量不足,且每年干旱程度与干旱出现的时期及持续时间有关。因此,深入了解冬小麦干旱和需水的生理生态特性是保证粮食高产稳产的重要研究课题。研究认为,高等植物对水分胁迫-复水的相应方式是胁迫解除之后存在短暂的快速生长以部分补偿胁迫造成的损失[3]。这种水分胁迫后复水引起的补偿生长效应可表现在形态结构、生理生化功能、产量和水分利用效率等方面[4~7]。目前的研究集中在作物关键生育期和部分器官方面,如根冠形态、生物量积累和产量,而对作物整体水平上的形态、光合产物分配格局及其动态的研究较少。文章通过探讨干旱和不同生育期复水对冬小麦光合产物分配格局的影响,旨在了解冬小麦水分调控下光合产物分配规律和动态以及干旱锻炼后复水生长补偿机制,为提高粮食生产和生物节水提供依据。
2 材料与方法
2.1 研究地概况
该实验在中国科学院封丘农业生态试验站进行,该站位于114 °24'E、35 °00'N,属半湿润偏旱地区,暖温带季风气候。年平均降水量为605 mm,年蒸发量为1 875 mm,其中3月、4月和5月总降雨量为101 mm,蒸发量为536 mm。年平均气温为13.9℃,≥0℃积温为5 100℃以上,无霜期为220 d左右。全年日照时数为2 300~2 500 h,日照率为55%。
2.2 栽培与管理
该研究在中国科学院封丘农田生态试验站防雨篷下进行。供试品种选择2个国审半冬性冬小麦品种,周麦18和济麦20。试验用聚氯乙烯(polyvinylchloride,PVC)管(与根系形态和根系垂直分布格局研究同步进行),直径为0.2 m,长为1.0 m。取封丘农田土,碾碎,过筛装管,每管装干土25 kg,土壤容重为1.35 g/cm3,田间最大持水量为26.2%(重量比)。施肥按尿素、磷酸二铵、硫酸钾分别为15 kg/hm2、15 kg/hm2和 10 kg/hm2进行,一次加入混匀装管,上留5 cm加入石英砂。播前该土壤含全氮 560 mg/kg、速 效 磷 8.90 mg/kg、速 效 钾232.80 mg/kg。2005年10月25日播种,每管9株,三叶期定苗。次年春返青期开始进行水分处理。共设4个水分处理,即W0(土壤水分含量保持在田间最大持水量的30% ~40%)、Wr1、Wr2、Wr3(分别为干旱处理后拔节期、孕穗期、开花期复水至田间持水量70% ~75%)。Wc(长期充足供水处理,保持在田间最大持水量的70% ~80%)为对照。表1为冬小麦不同生育期土壤水分重量含水量。在冬小麦孕穗期、开花期和收获期取样,每次每个品种破坏性取样3个PVC管。土壤水分含量采用烘干法测定,冬小麦生物量测定以单株统计植株根、茎、叶、鞘和穗的鲜重,105℃下15 min杀青,然后80℃下24 h烘至恒重,称干重。干样品密封保存,以备可溶性糖含量的测定。可溶性糖含量采用蒽酮比色法测定。光合产物分配指数(partitioning coefficient,PC)用植株各器官干重占总干重的比例表示。
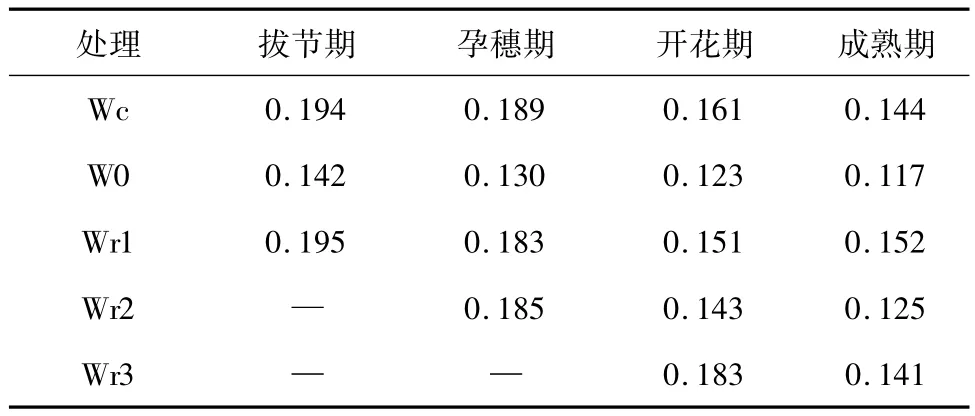
表1 冬小麦不同生育期土壤水分重量含量Table 1 Soil moisture content at different development stages of wheat
实验获得原始数据以冬小麦单株统计,文章中数据均为每个处理3个PVC管中27株冬小麦的平均值(±SE),采用SPSS软件对数据数进行方差分析、t-检验和相关分析。
3 结果
3.1 冬小麦生物量积累动态
作物生长对干旱后复水的滞后作用、复水后冬小麦形态变化和生长速度的变化需要一段时间后才能观察出来(见图1)。两个品种的 Wr1、Wr2和Wr3处理复水均表现出生长补偿,复水处理均高于W0处理,其中周麦18对复水反应敏感。Wr1和Wr2处理下,复水到下一生育期生物量表现出超补偿作用,高于对照Wc处理20% ~30%。Wr2复水处理下,生物量积累表现为周麦18复水后生物量积累比W0高35%,达到对照水平。济麦20生物量积累略高于干旱处理。开花期复水,作物复水后生物量补偿作用减小,周麦18和济麦20比干旱处理高10% ~15%,但均低于对照。拔节期、孕穗期复水能达到生物量积累的补偿作用。
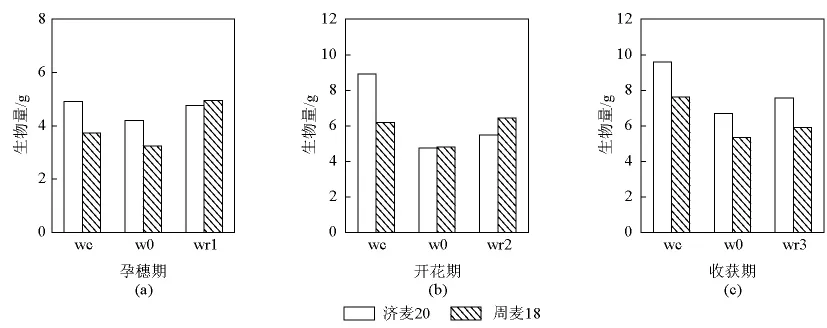
图1 不同生育期复水对冬小麦生长的影响Fig.1 Influence on growth of whole plant of rewatering at different developmental stages
3个复水处理均表现出生长补偿Wr>W0,其中Wr1增加幅度较大。济麦20和周麦18分别高于W0处理11.84%和34.43%,周麦18超补偿,高于Wc处理24.64%。Wr2处理济麦20和周麦18高于W0处理13.06%和25.03%,并且周麦18高于Wc处理3.91%。Wr3处理下,济麦20和周麦18分别高于W0处理11.41%和9.42%,周麦18在拔节期和孕穗期复水出现超补偿,其中拔节期复水补偿作用较强,济麦20在不同生育期复水补偿作用差别不大。
3.2 冬小麦器官水平光合产物分配
干旱后冬小麦不同生育期复水处理使冬小麦器官光合产物分配指数发生改变(见图2)。叶片光合产物分配指数在W0处理下均低于对照Wc处理,复水Wr1和Wr2处理叶分配指数有回升,但仍低于Wc处理。早期复水更有利于光合产物向叶片运输。
干旱胁迫使茎光合产物分配指数增加,W0处理茎分配指数高于Wc处理,Wr1复水处理后茎分配指数下降,周麦18和济麦20茎分配指数比W0处理下降了27.17%和6.60%,但高于Wc;Wr2、Wr3处理显示光合产物分配指数变化幅度逐渐减小,Wr2处理周麦18仍有20%下降,Wr3处理则无显著变化。干旱条件下叶鞘光合产物分配指数下降,不同时期复水具有回升,其中Wr1和Wr2有较显著的回升,Wr3回升较少。根的分配指数W0比Wc处理小,复水处理Wr1、Wr2和Wr3与W0差异不显著。早期干旱处理提高穗分配指数,孕穗期W0处理光合产物分配指数大于Wc,Wr1处理光合产物分配指数与Wc接近。开花期后干旱导致穗光合产物分配指数下降,W0小于Wc。复水处理Wr2和Wr3分配指数提高,与Wc无明显差异。干旱后冬小麦孕穗期和花期复水增加茎光合产物积累和灌浆期向穗中转运。

图2 不同复水处理下冬小麦光合产物分配指数Fig.2 The partitioning coefficients of wheat organ under different rewatering treatments
3.3 冬小麦不同器官可溶性糖含量
图3 (a)显示叶可溶性糖含量在长期干旱胁迫下,叶可溶性糖含量均高于Wc处理,其中开花期增高最多,达66%。Wr1叶可溶性糖含量与W0和Wc相比无显著差异,Wr2处理下济麦20和周麦18叶可溶性糖含量比W0下降32.99%和16.15%,但仍高于对照。干旱胁迫使大量糖滞留在叶中,随着胁迫解除叶片糖分物质迅速外运,叶可溶性糖含量下降。
干旱胁迫下冬小麦茎中临时库可溶性糖含量显著增加(见图3(b)),其中开花期茎可溶性糖含量比对照升高20% ~25%,两个品种间差异显著,周麦18高于济麦20。复水处理可使茎可溶性糖含量发生变化,Wr1处理茎可溶性糖含量与W0和Wc差异均不显著。Wr2处理茎中可溶性糖含量明显高于Wc处理,济麦20和周麦18分别高8.94%和20.12%。Wr3处理在收获期茎可溶性糖含量依然较高,开花期复水使灌浆期茎可溶性糖向穗的转运受到抑制。花前期特别是孕穗期适当的水分亏缺可促进可溶性糖在茎中的储存和向穗的转运。
根系可溶性糖含量对于土壤水分含量反映非常敏感(见图3(c))。干旱胁迫下根可溶性糖量高于Wc处理,其中孕穗期根可溶性糖含量差异最大,济麦20和周麦18根可溶性糖含量在W0处理下比Wc处理高93%和137%。Wr1、Wr2处理使根可溶性糖含量下降,低于Wc处理。相同处理下根可溶性糖含量表现为周麦18高于济麦20。拔节期和孕穗期复水使小麦根能通过改变渗透调节物质含量适应环境,尤其以拔节期复水调节能力强。

图3 冬小麦不同生育期复水可溶性糖含量Fig.3 WSC of winter wheat under different treatments
3.4 不同生育期复水对冬小麦产量的影响
小麦千粒重、穗粒数和穗数被称为小麦产量的3大要素,能够综合反映小麦生长发育过程中胁迫程度和光合产物在籽粒中的积累(见表2)。
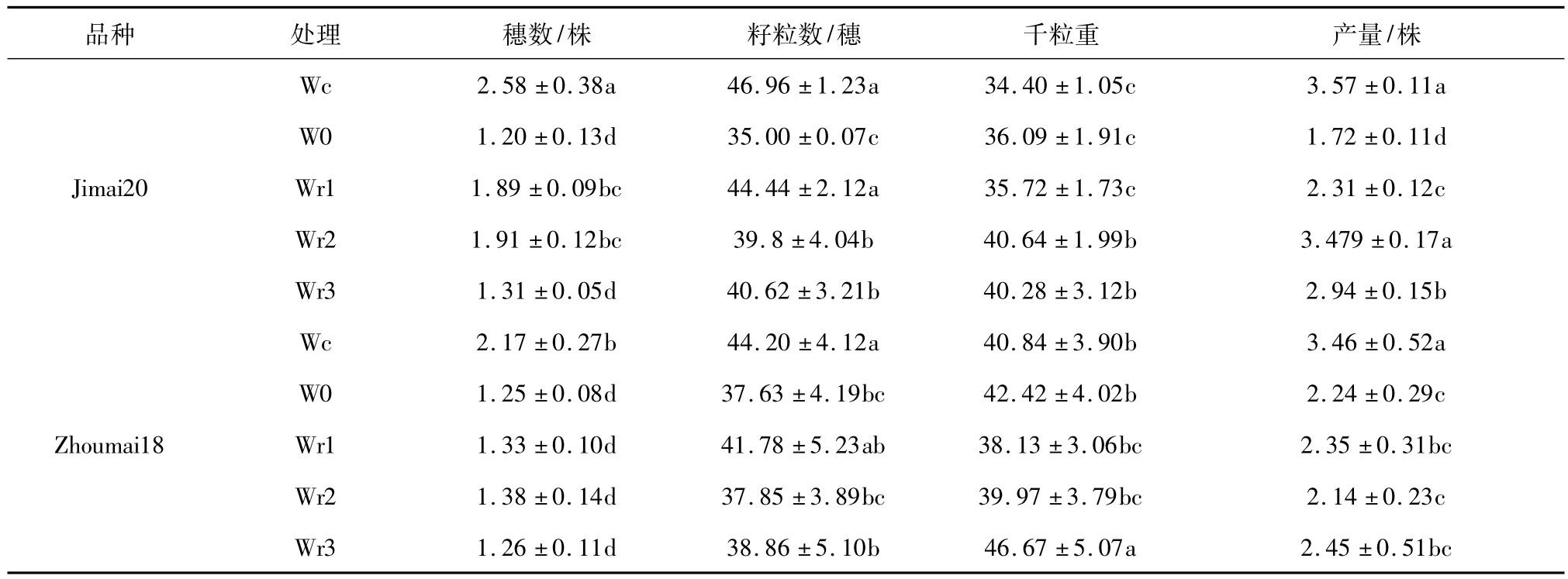
表2 干旱和复水对冬小麦产量因素的影响Table 2 Effects of drought and re-watering on yield factors of winter wheat
长期持续水分胁迫下和不同时期复水单株产量均比对照有显著下降,济麦20和周麦18分别减产51.95%和37.68%(P <0.05)。不同时期复水对产量补偿作用不同,济麦20和周麦18在Wr1、Wr2、Wr3下分别比对照减少 35.29%、30.53%、17.64%和 30.81% 、38.15%、29.19%。不同时期复水对产量的补偿途径不同,可分别从穗粒数和千粒重两个指标分析。
干旱胁迫冬小麦籽粒数下降,济麦20和周麦18分别比对照减少28.57%和14.86%。拔节期复水穗粒数减少幅度较小,济麦20和周麦18分别减少11.05%和2.39%。冬小麦拔节期开始生殖生长是生殖细胞四分裂的关键时期,拔节期灌水延缓拔节期生长时间能提高幼穗的形成和存活率,所以拔节水是维持冬小麦一定穗粒数的关键。籽粒千粒重代表籽粒的饱满程度,能反映小麦灌浆期灌浆速率受环境影响的程度。从表2可以看出W0处理下冬小麦千粒重略高于对照,济麦20和周麦18分别比对照高4.79%和6.47%,经检验没有达到显著水平。孕穗期和开花期复水处理下两个供试品种冬小麦千粒重提高了15.14% ~17.14%。籽粒光合产物来自于开花期期后即时光合产物和花前期临时库中储藏光合产物,开花期灌水能提高冬小麦旗叶的光合生产,并延缓衰老太高籽粒产量;花前干旱能促进临时库光合产物的储藏量,作物生育后期能较多转运到籽粒,所以开花期灌水能提高千粒重。
4 讨论
干旱胁迫导致冬小麦总生物量积累、叶面积等下降,改变光合产物在根冠间、器官间分配指数。干旱后复水冬小麦改变了光合产物分配格局。干旱胁迫下冬小麦减少光合产物向营养器官叶的分配,降低水分消耗[8,9],同时促进光合产物在茎中的储藏[10]。
干旱胁迫下,植物体内的可溶性糖大量积累,这与植物的抗旱性有关。作物分配一定量的可溶性糖用于渗透调节,这些渗透调节物质可以看做是一个与其他器官竞争的库,从整体水平上影响光合产物的积累、分配和利用[2]。叶片中可溶性糖是光合作用的初级产物,参与作物生长、代谢和抗逆[11],主要生理作用是参加渗透调节[12]。复水后叶片积累的可溶碳水化合物向外运输,提高叶片生产能力,在整株水平上表现出生长补偿。
干旱后,在花前期复水后茎可溶性糖含量仍然高于充足供水处理,茎可溶性糖含量的多少对花后同化物再动员有很大关系,茎可溶性糖含量可作为选育抗旱高产品种的重要指标。干旱胁迫总产量大幅度下降,不同时期复水均有不同程度产量补偿,拔节期复水主要提高小麦单株穗数,孕穗期和开花期复水以提高千粒重为主。不同品种产量补偿策略不同,遗传性状上济麦20比周麦18具有植株较高、分蘖力强、成穗率高的特点。济麦20主要提高成穗数,所以拔节水关键,而周麦18主要是提高灌浆期籽粒千粒重。胁迫下根可溶性糖含量增加说明根同化物分配提高,将升高根系活力和根干重[13]。胁迫下更多的碳水化合物贮存有利于耐旱和复水后的恢复,补偿作用与同化物分配密切相关,可溶性糖含量可能是一个比脯氨酸含量还好的标准。
总之,拔节期水分与幼穗的形成关系密切。拔节期复水能提高幼穗的存活率与穗粒数。开花期复水能促进后灌浆期作物生长和灌浆速率,对增加千粒重至关重要。
[1]Barnabás B,Jäger K,Fehér A.The effect of drought and heat stress on reproductive processes in cereals[J].Plant,Cell and Environment,2008,31(1):11 -38.
[2]谷艳芳,丁圣彦,高志英,等.干旱胁迫下冬小麦光合产物分配格局及其与产量的关系[J].生态学报,2010(5):1167-1173.
[3]Acevedo E,Hsiao T C,Henderson D W.Immediate and subsequent growth responses to maize leaves to changes in water status[J].Plant Physiology,1971,48:63l-636.
[4]罗远培,陈晓远.开花期复水对受旱冬小麦的补偿效应研究[J].作物学报,2001(4):512-516.
[5]陈晓远,罗远培.不同生育期复水对受旱冬小麦的补偿效应研究[J].中国生态农业学报,2002(1):35-37.
[6]袁永慧,邓西平.干旱与复水对小麦光合和产量的影响[J].西北植物学报,2004,24(7):1250-1254.
[7]赵丽英,邓西平,山 仑.开花前后变水条件对春小麦的补偿效应[J].应用与环境生物学报,2002,8(5):478-481.
[8]Blum A.Improving wheat grain filling under stress by stem reserve mobilization[J].Euphytica,1998,100(1):77 -83.
[9]Blum A,Belhassen E.Crop responses to drought and the interpretation of adaptation[J].Plant Growth Regulation,1996,20(2):135-148.
[10]Slafer G A,Abeledo L G,Miralles D J,et al.Photoperiod sensitivity during stem elongation phase as an avenue to rise potential yield in wheat[J].Euphytica,2001,119:191 -197.
[11]Robakidze E A,Bobkova K S.Carbohydrate accumulation in siberian spruce needles of various ages[J].Russian Journal of Plant Physiology,2003,50:573 -580.
[12]姜 东,于振文,李永庚,等.高产小麦营养器官临时贮存物质积运及其对粒重的贡献[J].作物学报,2003,29(1):31-36.
[13]Yim K O,Kwon Y W,Bayer D E.Growth responses and allocation of assimilates of rice seedlings by paclobutrazol and gibberellin treatment[J].Journal of Plant Growth Regulation,1997,16(1):35-41.
