甜菜M14品系短期盐胁迫下生理生化指标研究
2012-07-06贾姗姗马春泉陈思学李海英
贾姗姗,潘 钰,马春泉,于 冰,陈思学,李海英
(黑龙江大学a.生命科学学院;b.黑龙江省普通高等学校分子生物学重点实验室;c.农业微生物技术教育部工程研究中心,哈尔滨 150080)
0 引 言
土壤盐渍化问题一直是困扰农业生产的严重问题。由于我国人口多,可利用的耕地面积少,随着盐渍化问题的加重,粮食问题已成为我们现在面对的头等问题[1]。
植物在正常的生长条件下,植物本身就有可能产生活性氧等物质,如果外界环境没有明显的变化,植物自身的活性氧的产生与清除维持着一定的平衡,这时,植物不会受到活性氧等物质的伤害,可以正常生长发育[2]。如果植物所处的环境突然间发生变化,例如盐渍化等逆境,植物的内部环境的平衡就会被打破,这样活性氧的产生增多,而植物的清除机制却不能及时清除活性氧,最后导致植物的质膜受到活性氧的伤害,离子外渗,同时MDA含量增多,植物代谢紊乱,最终影响植物的生长乃至死亡[3-5]。
植物为了适应逆境,就会进化出一套植物耐盐机制,包括植物自身结构及器官功能特性对盐分的适应、合成渗透调节物质、离子调节、活性氧清除机制等方面。离子调节包括选择性吸收离子,或者将已经吸收细胞的离子外排出体外,亦或者将离子区域化于液泡或者老叶防止对植物新叶产生伤害;保护酶包括SOD、POD、CAT这3种酶相辅相成的来清除植物体内的活性氧;渗透调节物质包括吸收积累无机盐离子、合成积累有机小分子物质如脯氨酸等[6-8]。
野生白 花甜菜(Beta corolliflora Zoss.)具有良好的抗旱性,耐寒抗霜性,耐盐性和无融合生殖等优良特性。甜菜M14品系是二倍体栽培甜菜(Beta vulgaris L.)和四倍体野生白花甜菜(Beta corolliflora Zoss.)通过远缘杂交获得的带有野生白花甜菜第9号染色体的单体附加系。前期的研究工作表明,甜菜M14品系具有野生白花甜菜的一些优良性状,如抗逆性,可以忍受NaCl浓度500mmol/L[9-12]。本试验就是研究甜菜 M14品系在不同盐浓度胁迫下短期的生理生化反应的变化过程,以此来揭示植物的抗逆规律,为今后进一步研究甜菜M14品系的其他抗逆研究提供基础。
1 材料和方法
1.1 植物材料
甜菜 M14品系(Chromosome 9monosomic addition line of Beta corolliflora Zoss.in Beta vulgaris L.,VV+C9,2n=18+1),其染色体组成中除了包含18条栽培甜菜染色体外,还附加有白花甜菜第9号染色体。
1.2 甜菜培养与处理
挑选甜菜M14品系饱满的种子,播种于盛有蛭石的发芽盒中,7d以后将其移入内部盛有霍格兰营养液的黑色不透明的槽子中,以泡沫为支持物每孔3棵苗,当植株能够在新环境中正常生长时留取长势好的幼苗(保证每孔1颗苗),当植株长到7d,挑选长势一致的幼苗分到4孔的黑色不透明的盛有霍格兰营养液小槽子中准备进行胁迫处理。当植株生长稳定时每天加NaCl胁迫处理使其终浓度分别达到200、400mmol/L,以空白为对照,分别于1、2、3、5、7d进行采样保存,测植物的质膜透性、MDA含量、脯氨酸含量等物质。
1.3 质膜透性测定
称取甜菜M14品系叶片0.5g于洗净的小烧杯中,用ddH2O将其洗净,再加入15mL ddH2O振荡使叶片全部沉没,然后将其放在真空干燥箱中抽走叶片细胞间的空气使叶片完全萎蔫并浸泡在水中,取出冷却后常温下测电导率E1,煮沸10min后冷却至室温后测电导率E2,计算叶片相对电导率(%)=E1/E2。
1.4 MDA含量测定
称取甜菜 M14品系叶片0.5g,加入少量TCA进行研磨,最后定容至10mL,放入100℃水浴中煮沸10min,取出冷却,测量溶液450nm、532nm和600nm处的吸光度。计算MDA浓度C(μmol/L) =6.45(A532- A600) -0.56A450;MDA含量(μmol/g)=C(μmol/L)×上清液总体积(mL)/样品鲜重(g)×1000。
1.5 脯氨酸含量测定
称取甜菜M14品系叶片1g,加入少量80%的乙醇研磨,将研磨液转移至10mL管子中,使溶液终体积为10mL,封口膜封好于暗室中浸提24h,然后在有活性炭的滤纸上进行过滤,去除残渣,离心备用。取上清液2mL于管子中,加入2 mL冰醋酸和2mL茚三酮试剂,封口膜封住,沸水浴10min,冷却后测定515nm处的吸光度。计算:
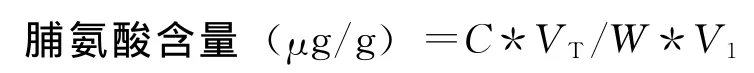
式中C为由标准曲线上查得的脯氨酸的质量,μg;VT为提取液总体积,mL;V1为测定液体积,mL;W 为样品质量,g。
1.6 可溶性蛋白质含量测定
称取甜菜M14品系叶片0.5g,加入5mL蒸馏水进行研磨,研磨至匀浆后转移至离心管中离心备用。吸取上清液1mL加入5mL考马斯亮蓝后摇匀放置10min,然后在595nm下比色,
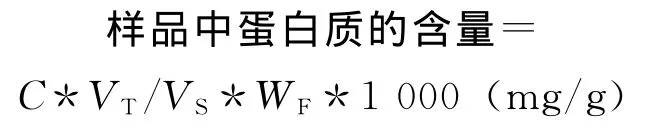
式中C为查得标准曲线值,μg;VT为提取液总体积,mL;WF为样品鲜重,g;VS为测定时加样量,mL。
2 结果与分析
2.1 甜菜M14品系叶片相对电导率的变化
甜菜M14品系叶片相对电导率的变化见图1。
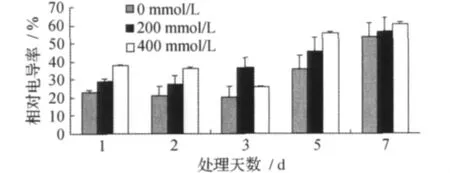
图1 叶片相对电导率Fig.1 Relative conductivity of leaves
由图1可见,随着NaCl处理时间的延长,0、200、400mmol/L NaCl胁迫处理下的甜菜M14品系的叶片相对电导率整体呈上升趋势,并且均在胁迫处理后的7d达到最高,在1、2、3、5dNaCl胁迫处理后的甜菜M14品系叶片的相对电导率与对照相比,在200、400mmol/L处理下相对比较高,而在7d胁迫处理后与对照相比,相差不大,趋于稳定。
2.2 甜菜M14品系叶片MDA含量的变化
甜菜M14品系叶片MDA含量的变化见图2。

图2 叶片MDA含量Fig.2 MDA content of leaves
由图2可见,随着NaCl胁迫处理时间的延长,与对照相比,甜菜 M14品系在200、400 mmol/L NaCl胁迫下的MDA含量随着时间的延长整体呈下降趋势,而对照相对来说是升高的;甜菜M14品系在200mmol/L NaCl胁迫处理下叶片的MDA含量在胁迫处理后的2d达到最低,而在400mmol/L NaCl胁迫处理下叶片的MDA含量在胁迫处理后的7d达到最低。
2.3 甜菜M14品系叶片脯氨酸含量的变化
甜菜M14品系叶片脯氨酸含量的变化见图3。
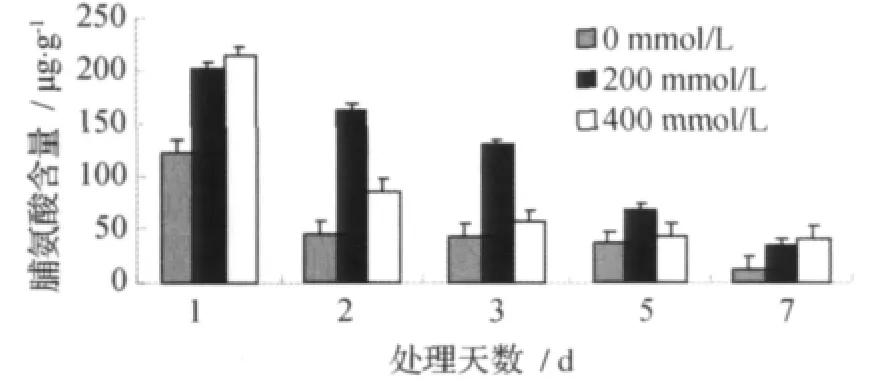
图3 叶片脯氨酸含量Fig.3 Proline content of leaves
由图3可见,甜菜M14品系叶片的脯氨酸含量随着不同浓度(0、200、400mmol/L)盐胁迫时间的延长整体呈下降趋势并且均在7d达到最低。
2.4 甜菜M14品系叶片可溶性蛋白质的变化
甜菜M14品系叶片可溶性蛋白质的变化见图4。
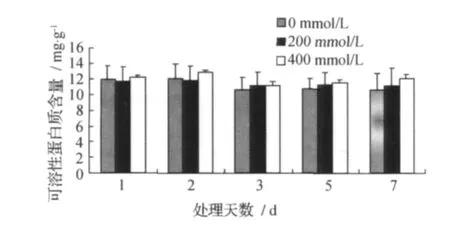
图4 叶片可溶性蛋白含量Fig.4 Soluble protein content
由图4可见,与对照相比,200、400mmol/L NaCl胁迫处理后的甜菜M14品系叶片的可溶性蛋白质含量随着盐胁迫时间的延长未有明显变化,可溶性蛋白质含量在400mmol/L NaCl胁迫处理后比对照组高些,而200mmol/L NaCl胁迫处理后的可溶性蛋白质含量在处理后的1、2d略低于对照组,在3、5、7d胁迫处理后略高于对照组。
3 讨 论
通过对试验结果分析得出,随着盐浓度的增大,甜菜M14品系叶片的电导率逐渐增大,细胞的质膜透性在盐胁迫下逐渐增大,但增加的幅度并不明显,而且同一浓度盐胁迫处理下随着时间的延长植物细胞的电导率也在逐渐增大,并且在处理后的7d达到最高,同时结合MDA含量随着盐浓度的增加及盐胁迫时间的延长,整体呈现下降趋势,也就是植物在逆境下迅速启动了保护系统来防止植物进一步被盐伤害;通过测定的脯氨酸和可溶性蛋白质含量可以看出,盐胁迫下最先响应的是脯氨酸,而可溶性蛋白质含量增幅不大。由此可以得出,甜菜M14品系可以在400mmol/L NaCl胁迫下生长良好。
[1]陈莎莎,兰海燕.植物对盐胁迫响应的信号转导途径[J].植物生理学报,2011,47(2):119-128.
[2]李春燕,陈思思,徐 雯,等.苗期低温胁迫对扬麦16叶片抗氧化酶和渗透调节物质的影响 [J].作物学报,2011,37(12):2293-2298.
[3]金光德,南桂仙.植物盐胁迫响应及耐盐的分子机制[J].农技服务,2011,28(10):1448-1449.
[4]孙天国,沙 伟.盐胁迫对甜瓜种子萌发及幼苗SOD、POD活性的影响 [J].安徽农业科学,2009,37(24):11509-11510.
[5]董兴月,林 浩,刘丽君,等.干旱胁迫对大豆生理指标的影响 [J].大豆科学,2011,30(1):83-88.
[6]吕 品,张 岩,李建华,等.植物细胞活性氧的产生和清除机制 [J].生物学教学,2010,35(2):4-5.
[7]马春泉,张 莹,崔 颖,等.甜菜 M14品系花期cDNA文库的构建及特意表达基因的筛选 [J].植物研究,2008,28(4):408-411.
[8]毛秀红,刘翠兰,燕丽萍.植物盐害机理及其应对盐胁迫的策略 [J].山东林业科技,2010,(4):129-130.
[9]Haiying Li,Hongxiang Cao,Yuguang Wang,et al.Proteomic analysis of sugar beet apomictic monosomic addition line M14 [J].Journal of Proteomics,2009,73:297-308.
[10]Chunquan Ma,Yuguang Wang,Lifa Wang,et al.I-dentification of a sugar beet BvM14-MADS box gene through differential gene expression analysis of monosomic addition line M14 [J].Journal of Plant Physiology,2011,168:1980-1986.
[11]李海英,马春泉,于 冰,等.利用mRNA差异显示技术分离甜菜M14品系特意表达基因的cDNA片段 [J].植物研究,2007,27(4):465-468.
[12]阮松林,童建新,赵杭苹.植物响应逆境胁迫蛋白质组学研究进展 [J].杭州农业科技,2007,(2):15-18.
