生活垃圾填埋土对乔灌木幼苗光合特性的影响1)
2012-06-28杨柳李君
杨 柳 李 君
(华东师范大学,上海,200062)
郑思俊 张庆费
(上海市园林科学研究所)
城市生活垃圾除小部分通过焚烧、堆肥处置外,大部分采用卫生填埋方式[1-2]处理。垃圾填埋不仅占用大量土地,浪费资源,而且垃圾的不断腐败分解使土壤营养元素、重金属、有毒物质等含量增加,土壤理化性质改变[3]。对已封场的填埋场进行生态恢复,实现填埋土资源化利用具有重要意义。目前对生活垃圾填埋场的研究多集中在土壤性质、污染处置、植被生理方面[4-6],填埋场生态恢复的工程设计、植被重建也有涉及[3,6],但垃圾填埋土对植物光合作用的影响方面研究甚少。笔者以老港生活垃圾填埋场不同封场期土壤为对象,研究了女贞、蚊母等6种乔灌木幼苗的光合、荧光特性,以期找到适合填埋场生态恢复的植物种类。
1 材料与方法
1.1 材料
试验在上海市园林科学研究所实验大棚内进行。供试植物为女贞(Ligustrum lucidum)、蚊母(Distylium racemosum)、香樟(Cinnamomum camphora)、红叶石楠(Photinia serrulata)、夹竹桃(Nerium oleander)、胡颓子(Elaeagnus pungens)6种喜光、抗性强常绿乔、灌木健康幼苗,其中胡颓子株高30~50 cm,其它5种植物株高100~150 cm。试验土采自上海老港生活填埋场封场于1995年、2000年、2005年的填埋单元,采土深度控制在距地面150(100)cm内。塑料盆规格为直径350 cm、高度560 cm,盆底有孔,盆下垫置直径500 cm和高度5 cm的塑料盘,用于回灌。
1.2 试验方法
2010年5月中旬,于3个填埋年限的垃圾土上分别种植红叶石楠、女贞等6种乔灌木的幼苗,每种植物各种4盆,共72盆。将所有盆栽置于大棚内,棚顶盖有塑料膜,用于遮雨。大棚四周通风,棚内外温差控制在±0.5℃以内。实时浇水,土壤相对湿度控制在40%~45%。
土壤测定:各期填埋土经自然干燥筛分后进行测定,土壤pH、EC值采用电位法,有机质采用重铬酸钾法,全氮采用半微量开氏法,全磷采用碱熔—钼锑抗比色法,水解氮采用碱解—扩散法,重金属采用盐酸—硝酸—高氯酸消解—ICP法。
光合参数测定:2011年8月中旬,选取晴朗无云的天气,于9:00—11:00将盆栽搬至室外空旷地后采用Li-6400(Li-Cor Inc.USA)光合仪对植株进行 Pn、Ci、Gs、Tr的测量。测量时,选取靠近植株顶端的第3~5片健康叶[7],每盆3片,每片叶待数据稳定后记录10个数据。
叶绿素荧光参数:采用Li-6400-40荧光叶室和Li-6400-01 CO2液化钢瓶对叶片PSⅡ最大光能转换效率 Fv/Fm、PSⅡ实际光化学量子效率ΦPSⅡ、光化学淬灭系数qP、非光化学淬灭系数qN进行测定。测定前将用于光合指标测定的叶片用暗适应叶片夹暗处理12 h,测量时饱和光强设定为500 μmol/m2·s,CO2摩尔分数设为 400 μmol·mol-1。
1.3 数据处理
数值运用 SPSS18.0统计软件进行 one-way ANOVA分析,选用LSD法进行多重比较,相关性分析采用Pearson法。
2 结果与分析
2.1 土壤因子
从表1可见,不同封场期填埋单元土壤养分因子差异显著,EC、全磷质量分数随填埋年限增大而减小,最小值为最大值的61.2%、62.2%;全氮年际变化与之相反;有机质、水解氮质量分数2000年垃圾土中最低,仅有71.37 g·kg-1、548.63 mg·kg-1,土壤pH值2000年垃圾土最高。总体上,各封场期土壤营养水平远超国家林业有机肥养分含量标准,其中水解氮超过标准5倍以上,可能对植物造成营养胁迫[8]。

表1 不同封场期垃圾填埋土养分指标
从表2可见,不同垃圾土重金属差异显著,其质量分数随填埋年限变化规律不明显,但除砷外均为2000年垃圾土最低,土壤砷质量分数是《土壤环境质量标准GB15618-1995》三级的5倍,锌为国家标准三级的两倍多,镉也为国家标准三级的一倍多。当重金属过量时将影响植物对水分和营养元素的吸收和运输,对叶绿素合成有关酶系统和电子传递产生负面影响,抑制其光合作用[9-11]。
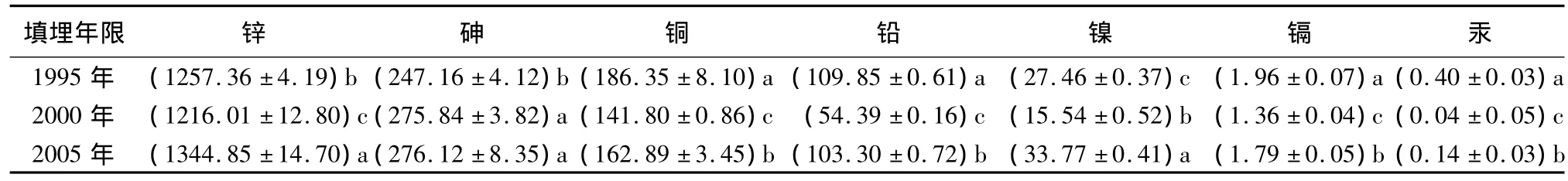
表2 不同封场期垃圾填埋土土壤重金属质量分数 mg·kg-1
2.2 不同填埋年限的垃圾土中各乔灌木的光合特征
表3表明,生长于1995、2000、2005年垃圾填埋土中的6种乔灌木幼苗,夹竹桃Pn最高,女贞、胡颓子、蚊母、香樟、石楠Pn均值分别为夹竹桃的73.9%、50.7%、49.5%、49.2%、43.0%。蚊母、石楠Pn随填埋年限变化趋势相近,1995年垃圾土中最高;女贞、胡颓子2000年垃圾土中Pn显著高于其他2期,随填埋龄先升后降;夹竹桃Pn变化趋势则随填埋龄先降后升,与香樟同为2005年垃圾土中最高。香樟、石楠的Gs在不同填埋龄垃圾土中差异不显著,胡颓子Gs为1995年填埋土中最高;女贞、蚊母Gs变化呈先降后升趋势,夹竹桃与之相反。Ci、Tr与Gs正相关,变化趋势与 Gs相似(rCi=0.744,PCi=0 <0.05;rTr=0.927,PTr=0 <0.05)。
2.3 不同填埋年限的垃圾土中各乔灌木的荧光特征
植物叶片荧光参数的变化可以有效地衡量植物的受害程度和光合潜能的高低[12],其中Fv/Fm是植物光合潜力的灵敏指标[13]。由表4可知,6种植物Fv/Fm最高值相近,最大光合作用潜力相当,但出现的填埋年限不同。女贞、石楠Fv/Fm随填埋年限减小,1995年垃圾土中最高;夹竹桃、胡颓子Fv/Fm随填埋年限增大,2005年垃圾土中最高;蚊母、香樟Fv/Fm变化呈先降后升趋势,2000年垃圾土中最低。各幼苗Fv/Fm值不同程度地低于0.83,说明植物处于受胁迫状态,光合作用受到不同程度的抑制[14]。夹竹桃ΦPSⅡ均值显著高于其他植物,女贞、胡颓子、香樟、蚊母、石楠ΦPSⅡ均值分别为夹竹桃的70.0%、66.1%、63.9%、42.3%、38.6%。不同填埋年限垃圾土中各植物 ΦPSⅡ差异显著:蚊母、石楠1995年垃圾土中最高,女贞、夹竹桃、胡颓子为2000年垃圾土中最高,香樟ΦPSⅡ最高值出现于2005年垃圾土。各试验幼苗qP与ΦPSⅡ变化趋势相似。女贞、胡颓子在不同填埋年限垃圾土中,qN差异不显著。

表3 不同封场期垃圾填埋土中各植物幼苗光合指标
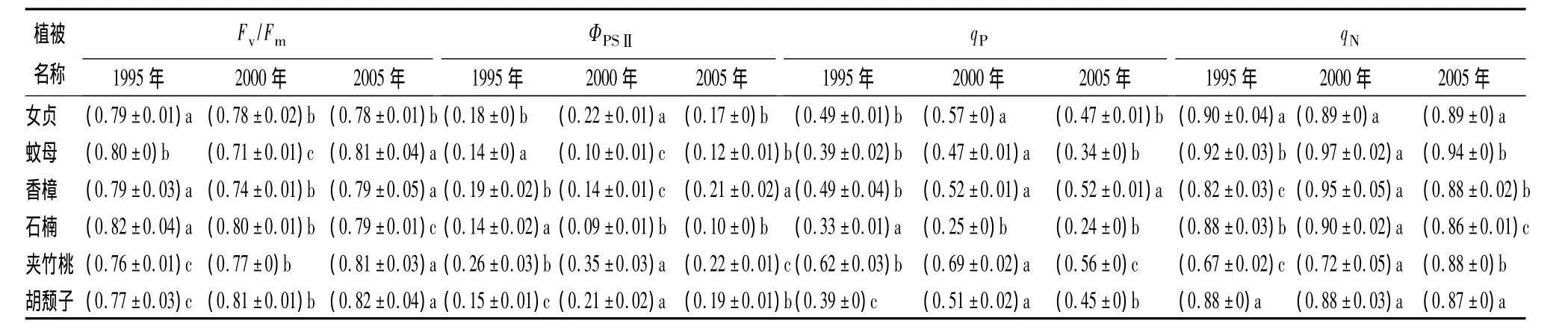
表4 不同封场期垃圾填埋土中各植物幼苗荧光指标
2.4 光合指标与土壤因子间的相关性
由表5可知:土壤因子与光合指标的相关性,pH、全氮、水解氮与Pn负相关,在一定程度上限制了光合作用的进行。Pn随土壤中砷、镉、铜、汞质量分数的升高显著减小,垃圾土中过量的砷、镉、铜、汞使得幼苗叶肉光合能力遭受限制[15],光合作用遭受胁迫。土壤因子与叶绿素荧光指标的相关性表现为,Fv/Fm与显著负相关,植物光合作用潜力受到pH、砷抑制。ΦPSⅡ与 EC、有机质等营养因子,全部所测重金属负相关,部分显著负相关。垃圾土中各因子,尤其是重金属的改变使得植物实际光化学效率下降。全磷、水解氮等营养元素,镉、镍等重金属与qP显著负相关,过量营养元素、重金属质量分数抑制了PSⅡ反应中心“开放”程度,使得QA重新氧化量减小。qN与土壤因子的相关性与qP相似,受土壤因子的影响与qP相似。
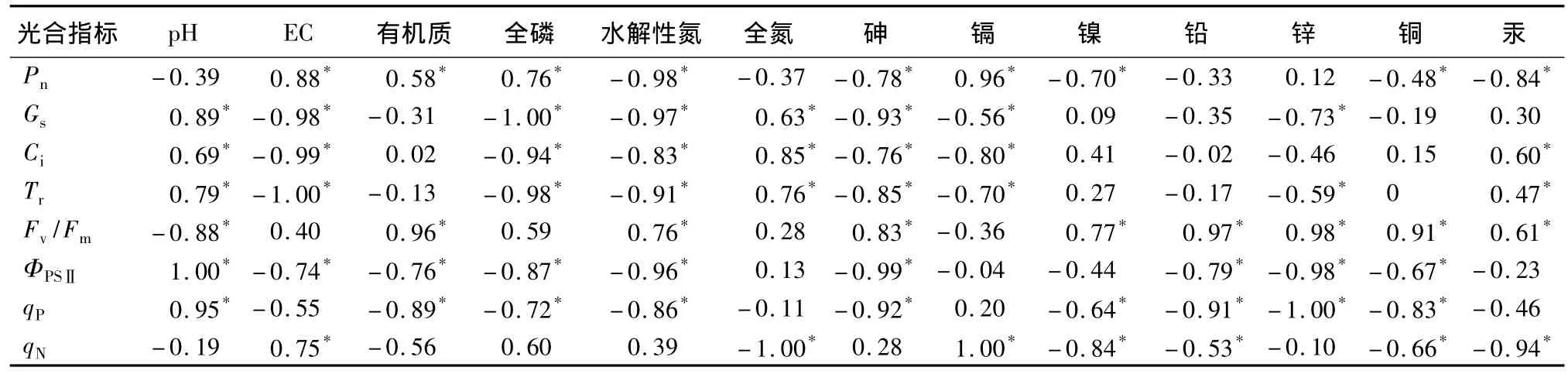
表5 光合指标与土壤化学因子、土壤重金属间的相关系数
3 结论与讨论
3.1 垃圾土中各土壤指标对乔灌木光合特征的影响
Pn下降的原因分气孔限制和非气孔限制[15]:Pn与Ci变化趋势一致时为气孔限制,否则为非气孔限制[16]。本实验不同填埋年限垃圾土中6种幼苗 Pn与 Ci显著负相关(r=-0.473,P=0.048<0.05),两者变化趋势相反,为非气孔限制。垃圾填埋土营养水平、重金属含量高,各种有害物质掺杂在一起,可能对植物光合造成复合抑制[17],对土壤因子与光合指标进行相关性分析:pH、全氮、水解氮与Pn负相关,在一定程度上限制了光合作用的进行。Pn随土壤中砷、镉、铜、汞质量分数的升高显著减小,说明垃圾土中过量的砷、镉、铜、汞使得幼苗叶肉光合能力遭受限制[18],光合作用遭受胁迫。结合图1~图4看出6种植物光合均受抑制,叶肉细胞受到不同程度损害,其中蚊母Gs、Ci是6种幼苗中最大的,Pn却相对较小,说明其叶肉细胞受到损害最大,光合受到的抑制最强。女贞、夹竹桃Gs较小,而Pn较大,光合作用受到的抑制相对较小。净光合速率反映了立地质量状况[19],就3种不同填埋年限实验垃圾土而言:蚊母、石楠在1995年垃圾土中Pn最大,1995年垃圾土对蚊母、石楠光合作用相对有利;2000填埋垃圾土立地条件最有利于女贞、胡颓光合能力的发挥;夹竹桃、胡颓子则在2005年垃圾土中光合能力最强。
3.2 垃圾土中各土壤指标对乔灌木荧光特征的影响
Fv/Fm随pH、砷升高而下降,说明在高pH、高砷胁迫下,叶片发生了光抑制或PSⅡ复合体受损害,植物光合作用潜力下降[20]。ΦPSⅡ是光下 PSⅡ反应中心部分关闭情况下的实际光化学效率,反映了PSⅡ的光能转换效率[21]。夹竹桃 ΦPSⅡ最高,实际光化学效率最大,其次为胡颓子、女贞、香樟,蚊母和石楠实际光化学效率相对最低,对光能的利用率最低。随着土壤肥力的增加、重金属浓度的增大,植物ΦPSⅡ不断下降,说明高营养水平、高重金属浓度抑状态直接相关[22]。qP与PSⅡ反应中心“开放”状态[23],PSⅡ稳定原初电子受体QA的氧化还原状态有关;qN表示吸收的光能以热的形式耗散掉的部分,是植物保护PSⅡ的重要机制[24]。本研究下全磷、水解氮等营养元素,镉、镍等重金属与qP显著负相关,说明过量营养元素、重金属含量抑制了PSⅡ反应中心“开放”程度,使得QA重新氧化量减小。qP降低时,qN并未相应升高,说明在非光胁迫条件下光合机构遭受破坏,光合受到抑制[25]。6种幼苗中夹竹桃qP最高,光化学潜能最大[26],女贞、香樟、胡颓子次之,蚊母、石楠最小。各植物幼苗qN相近,热耗散能力相当,对光合结构的保护能力相当。本试验条件下,相对其他垃圾土,蚊母、石楠在1995年垃圾土中光合潜力大,实际光合能力较高;女贞、胡颓子在2000年垃圾土中PSⅡ开放状态好,光能利用率高,光合机构损害小;香樟在2005年垃圾土中光合潜能高,实际光合效率、有效光合效率强;夹竹桃在2005年垃圾土中潜在光合能力强,但实际光合能力并不是最大。
综上所述,垃圾土中水解氮、砷超过国家标准5倍以上,锌为国标三级的1倍多,成分复杂的垃圾土中多种有害物质复合胁迫对乔灌木幼苗光合、荧光造成不同程度的抑制:6种植物中夹竹桃光合、荧光受到的抑制最小;蚊母、石楠对垃圾土环境敏感,光合作用受到的抑制相对最大。不同植物在不同垃圾土中光合能力差异显著,相对其他期土壤,蚊母、石楠光合能力在碱度相对较大,营养水平及铜、铅、镉、汞含量较高的1995年垃圾土中较强;女贞、胡颓子在营养水平中等、重金属质量分数相对最小2000年垃圾土中光合能力较好;营养水平低,锌、砷、镍质量分数高的2005年垃圾土能使夹竹桃、香樟光合能力得到最大水平的发挥。
[1]胡斌,丁颖,吴伟祥,等.垃圾填埋场恶臭污染与控制研究进展[J].应用生态学报,2010,21(3):785-790.
[2]刘艳辉,魏天兴,孙毅.城市垃圾填埋场植被恢复研究进展[J].水土保持研究,2007,14(2):108-111.
[3]李俊,舒为群,陈济安,等.垃圾填埋场土壤酶活性与化学性质和微生物数量的关系研究[J].生态学杂志,2005,24(9):1043-1047.
[4]郭亚丽,赵由才,徐迪民.上海老港生活垃圾填埋场陈垃圾的基本特性研究[J].上海环境科学,2002,21(11):669-672.
[5]邹龙生,唐婧,叶凯贞.上海老港垃圾填埋场渗滤液处理工艺的选择[J].云南民族大学学报:自然科学版,2011,21(1):23-26.
[6]温馨,封莉,王辉,等.生活垃圾填埋场不同封场期场地植物抗氧化酶活性[J].生态学杂志,2010,29(8):1612-1617.
[7]Theodore T K,Paul J K,Stephen G P.The physiological ecology of woody plants[M].New York:Academic Press Inc,1991:15-40.
[8]曾琳,王更亮,王广东.氮磷钾营养水平对观赏向日葵生长发育及光合特性的影响[J].西北植物学报,2010,3(6):1180-1185.
[9]Clemens S,Palmgren M G,Kramer U.Along way ahead:Understanding and engineering plant metal accumulation[J].Trends in Plant Science,2002,7(7):309-315.
[10]Krupa Z.Cadmium against higher plant photosynthesis:a variety of effects and where do they possibly come from?[J].Zeitschrift für Naturforschung C,1999,54:723-729.
[11]Lin R Z,Wang X R,Luo Y,et al.Effects of soil cadmium on growth,oxidative stress and antioxidant system in wheat seedlings(Triticum aestivum L.)[J].Chemosphere,2007,69:89-98.
[12]Llndgen K,Hallgren J E.Cold acclimation of Pinus contorta and Pinus sylvestris assessed by chlorophyll fluorescence[J].Tree Physiology,1993,13:97-106.
[13]张黎萍,荆奇,戴廷波,等.温度和光照强度对不同品质类型小麦旗叶光合特性和衰老的影响[J].应用生态学报,2008,19(2):311-316.
[14]Demming-Adams B,AdamsⅢ WW.Xanthophyll cycle and light stress in nature:Uniform response to excess direct sunlight among higher plant species[J].Planta,1996,198:460-470.
[15]Schützendübel A,Schwanz P,Teichmann T,et al.Cadmium-induced changes in antioxidative systems,hydrogen peroxide content,and differentiation in Scots pine roots[J].Plant Physiol,2001,127(3):887-898.
[16]Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Annual Review of Plant Physiology,1982,33:317-345.
[17]白莉萍,宋金洪,辛涛,等.施用城市污泥对小叶黄杨光合特性和生长的影响[J].应用生态学报,2010,21(4):1026-1030.
[18]Schröder W P,Arellano J B,Bittner T,et al.Flash-induced absorption spectroscopy studies of copper interaction of with photosystem Ⅱ in higher plants[J].J Biol chem,1994,269(52):32865-32870.
[19]冯玉龙,李继武,王文章,等.用净光合速率评价森林立地质量的可行性[J].东北林业大学学报,1996,2(6):80-84.
[20]梁文斌,薛生国,沈吉红.锰胁迫对垂序商陆光合特性及叶绿素荧光参数的影响[J].生态学报,2010,30(3):619-625.
[21]李晓,冯伟,曾晓春.叶绿素荧光分析技术及应用进展[J].西北植物学报,2006,26(10):2186-2196.
[22]Wang K F,Xu C X,Zhao F H,et al.The effects of water stress on some in vivo chlorophyll a fluorescence parameter soft wheat flag leaves[J].Acta Biophysica Sinica,1997,13(2):273-278.
[23]许大全.植物光胁迫研究中的几个问题[J].植物生理学通讯,2003,39(5):493-495.
[24]Binder W D,Fielder P.Chlorophyll fluorescence as in indicator of frost hardiness in white spruce seedlings from different latitudes[J].New Forests,1996,11:233-253.
[25]师生波,李和平,王学英,等.高山植物唐古特山莨菪和唐古特大黄对强太阳辐射光能的利用和耗散特性[J].植物生态学报,2007,1:133-141.
[26]Ehlert B,Hinvha D K.Chlorophyll fluorescence imaging accurately quantifies freezing damage and cold acclimation responses in a Aabidopsis leaves[J].Plant Methods,2008,4:1-7.
