盐碱胁迫对角果碱蓬幼苗光合特性的影响1)
2012-06-13庞秋颖张爱琴阎秀峰
魏 磊 庞秋颖 张爱琴 国 静 阎秀峰
(东北油田盐碱植被恢复与重建教育部重点实验室(东北林业大学),哈尔滨,150040)
人类不断开发和不合理利用土地,使土壤盐碱化程度不断加重,并成为威胁农业生产的主要逆境胁迫之一[1]。土壤中的致害盐类除了以NaCl为主的中性盐以外,还有以Na2CO3和NaHCO3为主的碱性盐[2]。碱性盐对植物不仅有与中性盐类似的离子毒害和渗透胁迫作用,还有高pH值及显著降低矿质元素的利用率等危害[3]。盐害对光合作用的抑制是限制植物生长的主要因素[4-5]。已有研究表明[6],NaCl胁迫通过多个方面抑制植物的光合作用,包括影响植物对CO2的吸收、运输及固定,影响植物对光能的吸收、转化和电子传递以及抑制原初产物的利用、转运和分配。然而,到目前为止,碱性盐对植物光合作用影响的研究相对较少[7-9]。
碱蓬属(SuaedaForsk.ex Scop.)植物为高耐盐真盐生植物,是研究植物耐盐碱机制的良好材料。目前对碱蓬属耐盐碱机制的研究多以盐地碱蓬(S.salsa)为材料[10-11]。角果碱蓬(S.corniculata)为碱蓬属1年生草本,主要生长在我国东北的松嫩盐碱荒地,具有很强的耐盐碱能力,却一直被人们忽视。本文选取角果碱蓬为材料,利用中性盐(NaCl)和碱性盐(NaHCO3)对角果碱蓬幼苗进行胁迫处理,分析比较不同盐浓度下两种盐对其光合生理特性和叶绿素荧光的影响,为进一步研究角果碱蓬响应盐碱胁迫的生理机制提供理论依据。
1 试验材料
角果碱蓬种子采集自黑龙江省肇东市周边盐碱地区,挑选籽粒饱满的角果碱蓬黑色种子,在水中浸泡2 h,播种于盛有洗净河沙的塑料盆中,温室培养,每天浇灌适量的1/2Hoagland营养液(蒸馏水配制,pH=6.2±0.1)。待长至4周后,将幼苗转移至水培盆中,每天更换1/2Hoagland营养液,培养1周后分别用含有 100、200、300、400 mmol·L-1NaCl和 50、100、150、200 mmol·L-1NaHCO3的 1/2Hoagland 营养液处理3 d,以1/2Hoagland营养液正常培养生长的幼苗作为对照。
2 试验方法
植株鲜质量及干质量的测定 将胁迫处理后的植株用去离子水冲洗干净,吸干表面水分,称量全株鲜质量。然后放入烘箱中105℃杀青10 min,再以80℃烘至恒定质量,称干质量。处理和对照每组5个重复。
光合参数测定 选择9:00—11:00时段进行光合参数测定,使用LI—6400便携式光合仪(LI-Cor,Inc,美国)测定角果碱蓬幼苗在光强1 000 μmol·m-2·s-1下的净光合速率、蒸腾速率、气孔导度、胞间CO2摩尔分数等指标。对照和处理每组各测定5株幼苗。
叶绿素荧光参数的测定 叶绿素荧光参数用英国Hansatech公司生产的FMS—2便携式脉冲调制荧光仪进行测定。先将叶片用暗适应夹进行30 min的暗适应,然后测定初始荧光(F0)、最大光化学效率(Fv/Fm)、光化学淬灭系数(qP)和电子传递速率(RE,T)等。每组进行5次重复测量。
数据处理 试验数据使用Microsoft Excel和SPSS PASW Statistics v18.0软件进行分析统计,并利用单因素方差分析(one-way ANOVA)比较不同数据组间的差异。
3 结果与分析
3.1 盐碱胁迫对角果碱蓬幼苗鲜质量及干质量的影响
如表1所示,用NaCl和NaHCO3分别处理角果碱蓬幼苗时,随着中性盐(NaCl)和碱性盐(NaHCO3)处理浓度的增大,角果碱蓬幼苗的鲜质量、干质量与盐浓度总体上均成负相关。
NaCl处理下,当其浓度为 100 mmol·L-1时,处理组的鲜质量与对照组的鲜质量差异不显著,但当NaCl≥200 mmol·L-1后,随着盐浓度的增大,处理组鲜质量值明显减小。当400 mmol·L-1NaCl的高盐处理时,处理组的鲜质量值比对照组减少了55.7%,植株出现萎蔫现象。而当NaHCO3处理时,在50~150 mmol·L-1浓度范围内,处理组的鲜质量与对照组相比有所减小,但差异不显著,而用200 mmol·L-1NaHCO3处理时,处理组植株的平均鲜质量减少了61.7%,植株生长出现严重萎蔫,趋于死亡。
与鲜质量的变化情况相比,在NaCl、NaHCO3分别处理后,幼苗的干质量变化均不明显。与对照相比,当 NaCl≤300 mmol·L-1或 NaHCO3≤150 mmol·L-1时,处理组的干质量值均逐渐减小,但下降幅度较小,在最高浓度 400 mmol·L-1NaCl或 200 mmol·L-1NaHCO3时,此两种高浓度处理组的干质量分别下降了39.1%和31.9%。
可见,角果碱蓬有一定的耐盐碱能力,当盐碱胁迫浓度在 NaCl≤300 mmol·L-1或 NaHCO3≤150 mmol·L-1范围时,植株可维持相对正常生长,而当盐碱浓度达到 400 mmol·L-1NaCl或 200 mmol·L-1NaHCO3时,对植株的伤害较大,影响了角果碱蓬幼苗的正常生长。
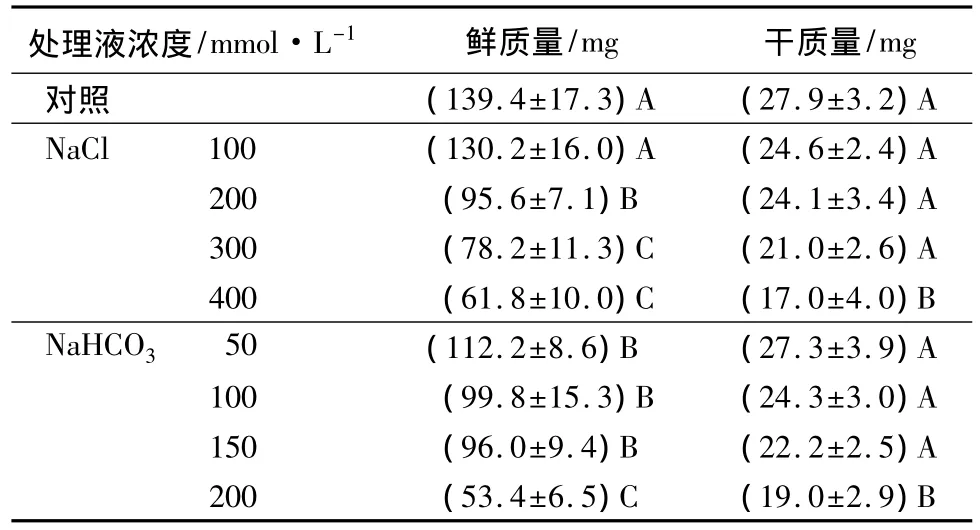
表1 盐碱胁迫对角果碱蓬幼苗鲜质量及干质量的影响
3.2 盐碱胁迫对角果碱蓬幼苗光合参数的影响
3.2.1 盐碱胁迫对角果碱蓬幼苗净光合速率、蒸腾速率的影响
如表2所示,当盐碱处理浓度分别在NaCl≤200 mmol·L-1或 NaHCO3≤100 mmol·L-1范围时,角果碱蓬幼苗净光合速率变化均不显著,但随着盐碱浓度的增大,幼苗的净光合速率均迅速下降,当达到最大盐碱处理浓度 400 mmol·L-1NaCl或 200 mmol·L-1NaHCO3时,与对照组相比,处理组的净光合速率值分别减少了73.7%和76%。两种盐处理对蒸腾速率的影响规律差异较大。与对照相比,在NaCl处理组中,蒸腾速率随盐浓度的增大出现缓慢下降趋势,而NaHCO3处理组中,随着NaHCO3浓度的增大,蒸腾速率出现先升高后降低的趋势,且在低浓度 NaHCO3处理(50、100 mmol·L-1)条件下,此两种浓度的处理组蒸腾速率与对照组相比分别升高 56.5%和 13.6%,而当 NaHCO3≥150 mmol·L-1后,处理组的蒸腾速率显著低于对照组。表明角果碱蓬的光合作用效率易受到盐碱胁迫的影响,并且在有碱性盐(NaHCO3)的胁迫下,其光合作用受影响程度更大。
3.2.2 盐碱胁迫对角果碱蓬幼苗气孔导度、胞间CO2摩尔分数的影响
从表2中可以看出,在NaCl、NaHCO3分别处理幼苗后,与对照相比,随着盐碱浓度的增大,此两种盐处理下的幼苗气孔导度均显著下降。400 mmol·L-1NaCl或 200 mmol·L-1NaHCO3胁迫时,这两个处理组的气孔导度分别下降了88.7%和89%。而NaCl、NaHCO3分别处理后,幼苗的胞间CO2摩尔分数与对照相比,随盐碱浓度的增大均有轻微上升趋势,但变化并不显著。表明盐碱胁迫对角果碱蓬幼苗的气孔伤害较为严重,随着浓度的增大,气孔开度减小,进而限制了蒸腾作用,使得蒸腾速率呈现下降趋势。在重度盐碱逆境下,叶肉细胞可能受到伤害,碳同化受到抑制,气孔与外界交换气体的作用受限,外界CO2进入量减少,使得胞间存留了一定量的CO2,因此盐碱胁迫后期胞间CO2摩尔分数呈现微升高。
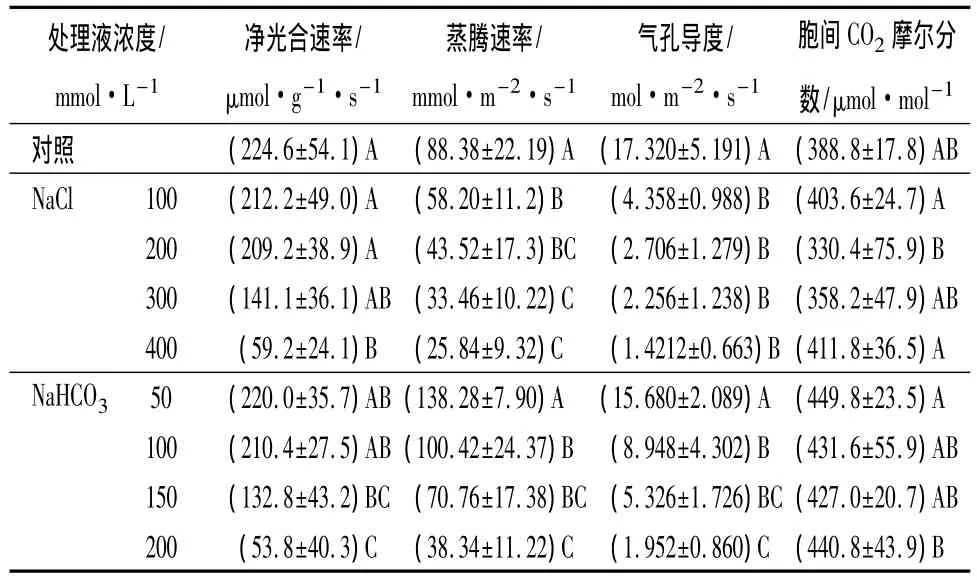
表2 盐碱胁迫对角果碱蓬幼苗光合参数的影响
3.3 盐碱胁迫对角果碱蓬幼苗叶绿素荧光特性的影响
叶绿素荧光各项参数被称为研究光合作用的重要探针,反映光能的利用情况。初始荧光(F0)是在已经暗适应的条件下,检测光系统Ⅱ(PSII)反应中心处于完全开放时的荧光产量,与叶绿素浓度有关[12]。从表3中可以看出,与对照相比,随着盐碱胁迫浓度的增大,NaCl或NaHCO3处理的F0逐渐增大。与NaCl胁迫相比,NaHCO3胁迫下F0值升高趋势更明显,这表明,在盐碱胁迫下,PSII反应中心受到了严重破坏,且碱性盐(NaHCO3)胁迫对于角果碱蓬幼苗PSII的伤害更为严重。
Fv/Fm是可变荧光与最大荧光的比值,表示PSII最大光化学量子产量。与对照相比,各处理组的Fv/Fm变化趋势与盐碱浓度成负相关,其变化幅度较小,最高浓度处理组(400 mmol·L-1NaCl或200 mmol·L-1NaHCO3)均下降了 10%左右(表 3),表明在盐碱逆境胁迫的条件下,光合作用的PSII反应中心受到强光的影响,并发生光抑制现象。
光化学淬灭系数(qP)反映PSII吸收的光能在光化学反应电子传递中的份额,并在一定程度上反映PSII反应中心的开放程度。与对照相比,随盐碱浓度的增大,各处理的qP值逐渐减小(表3),表明用于光化学反应的光能逐渐减少,并且PSII反应中心的电子传递受到了抑制。
PSII非循环光合电子传递速率(RE,T)是反映实际光强条件下的表观电子传递效率,体现PSII反应中心电子传递情况,用单位时间电子传递过程所吸收光量子数目来表示[13]。与对照相比,随着盐碱浓度的增大,各处理的RE,T逐渐减小。与NaCl处理比,NaHCO3碱性盐胁迫的各处理RE,T受影响更大(表3),表明盐碱胁迫影响了PSII反应中心电子传递效率,降低了光合电子的传递能力,且NaHCO3胁迫对植株的影响更显著。
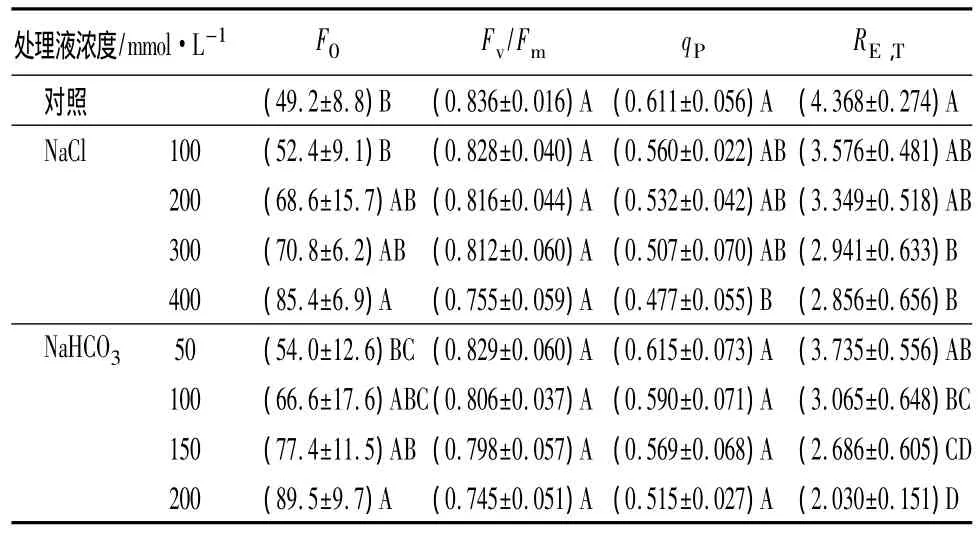
表3 盐碱胁迫对角果碱蓬幼苗叶绿素荧光参数的影响
4 讨论
植物鲜质量值及干质量值的变化能较好地反映植物在受到逆境胁迫时生物量的变化情况,尽管生物量是一个综合指标,受到多种因素的影响[14],不能完全反映植物受逆境胁迫的情况,但盐胁迫下生物量的变化可作为评估盐胁迫程度和植物抗盐能力的指标之一。本研究表明,在NaCl或NaHCO3胁迫下,低浓度盐碱处理时,角果碱蓬幼苗的地上部分鲜质量下降均比较缓慢,但高浓度盐碱胁迫下,鲜质量值降低的幅度均明显增大。表明角果碱蓬具有一定的耐盐能力,但 400 mmol·L-1NaCl或 200 mmol·L-1NaHCO3分别作用时,均能够破坏幼苗的正常生理代谢活动,对植株的伤害较大。
光合作用是生物稳定收集可用能量的生产行为。净光合速率可直接体现光合系统功能,可作为衡量植株系统工作正常与否的重要指标[15]。光合速率下降分为气孔限制和非气孔限制。采用Farquhar等[16]提出的光合作用气体交换模型,可判断气孔导度是否限制了光合作用速率。吴成龙等[17]在研究菊芋耐碱性时发现:在碱胁迫下产生的高pH值毒害降低了叶肉细胞的光合活性,表明非气孔限制是菊芋光合速率降低的主要原因。崔强等[18]在NaHCO3胁迫金银忍冬苗木时发现:在幼苗发育的不同时期影响光合作用下降的因素可能不同,在短期轻度胁迫时导致光合下降的主导因素为气孔限制,而在长期轻度胁迫下则开始出现非气孔限制。本研究通过分析角果碱蓬幼苗在受到盐碱胁迫时各光合参数的变化情况发现:在一定盐碱浓度下(在200 mmol·L-1NaCl或 100 mmol·L-1NaHCO3分别作用时),幼苗可维持光合系统功能相对正常,对外界离子毒害和碱性环境有一定的抵抗能力,但随着胁迫浓度的增大,幼苗受损伤程度加剧,光合能力下降,难以维持正常的生理代谢。并且光合速率的降低并未伴随有胞间CO2摩尔分数的提高,胁迫很可能损坏了叶肉细胞,使其光合活性减弱,非气孔因素的作用可能是限制角果碱蓬幼苗光合作用的重要因素。
叶绿素荧光动力学技术是测定叶片光合功能的快速、无损伤的重要方法,在测定叶片光合作用过程中,光系统对光能的吸收、传递、耗散、分配等方面具有独特的作用,与“表观性”的气体交换指标相比,叶绿素荧光参数更能反映“内在性”特点[19]。秦红艳等[20]通过研究盐胁迫下不同葡萄品种叶片各项荧光参数发现,盐胁迫可抑制葡萄叶片PSII的光化学活性,使PSII的原初光能转化效率、PSII潜在活性、PSII潜在光合作用活力受到抑制,同时还抑制了光合电子传递和光合膜的能量化作用。葛江丽等[21]在研究甜高粱盐胁迫下的荧光值变化时发现,50、100 mmol·L-1NaCl胁迫不会对甜高粱 PSII反应中心造成损坏,但200 mmol·L-1NaCl处理影响了甜高粱反应中心的活性。本研究发现,随着NaCl或NaHCO3浓度的增大,角果碱蓬幼苗F0逐渐增大,Fv/Fm、qP、RE,T逐渐减小,表明盐碱胁迫产生的离子毒害和高pH值对角果碱蓬幼苗的光合系统PSII反应中心造成了损伤,并影响了光合电子的传递,抑制了光系统Ⅱ的光合作用活力。
耐盐植物有较强的区域化能力,能把进入到细胞的离子积累在液泡中。高浓度的Na+和Cl-对这类植物的影响不大[22]。而当NaHCO3胁迫时,不仅会对植物产生离子毒害,还会给植物造成碱性的胁迫环境。在本试验中,当胁迫的Na+浓度一致时,HCO产生的碱性环境对角果碱蓬幼苗造成了更严重的影响,净光合速率、蒸腾速率及各荧光参数都有显著变化。HCO产生的高pH值环境使幼苗气孔受损,损坏了叶肉细胞,严重限制了植物的蒸腾作用,并且降低了光合电子的传递能力,破坏了PSII反应中心。与NaCl相比,NaHCO3胁迫对角果碱蓬幼苗光合作用的影响更为显著。
[1]赵可夫.盐生植物资源及盐碱土改良利用研究动态[J].资源与环境,1989,1(1):40-43.
[2]曲元刚,赵可夫.NaCl和Na2CO3对玉米生长和生理胁迫效应的比较研究[J].作物学报,2004,30(4):334-341.
[3]Wang Yucheng,Ma Hui,Liu Guifeng,et al.Analysis of gene expression profile ofLimonium bicolorunder NaHCO3stress using cDNA microarray[J].Plant Mol Biol Rep,2008,26(3):241 -254.
[4]Parida A K,Das A B.Salt tolerance and salinity effects on plants:a review[J].Ecotoxicology and Environmental Safety,2004,60:324-349.
[5]Tiwari B S,Bose A,Ghosh B.Photosynthesis in rice under a salt stress[J].Photosynthetica,1997,34(2):303-306.
[6]张娟,姜闯道,平吉成.盐胁迫对植物光合作用影响的研究进展[J].农业科学研究,2008,29(3):74-80.
[7]Chaves M M,Flexas J,Pinheiro C.Photosynthesis under drought and salt stress:regulation mechanisms from whole plant to cell[J].Annals of Botany,2009,103:551-560.
[8]Sudhir P,Murthy S D S.Effects of salt stress on basic processes of photosynthesis[J].Photosynthetica,2004,42(4):481-486.
[9]Shi L X,Guo J X.Changes in photosynthetic and growth characteristics ofLeymus chinensiscommunity along the retrogression on the Songnen grassland in northeastern China[J].Photosynthetica,2006,44(4):542-547.
[10]Wang Baoshan,Lüttge U,Ratajczak R.Effects of salt treatment and osmotic stress on V-ATPase and V-PPase in leaves of the halophyteSuaeda salsa[J].Journal of Experimental Botany,2001,52:2355-2365.
[11]Qiu Nianwei,Chen Min,Guo Jianrong,et al.Coordinate upregulation of V-H+-ATPase and vacuolar Na+/H+antiporter as a response to NaCl treatment in a C3halophyteSuaeda salsa[J].Plant Science,2007,172(6):1218-1225.
[12]张守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报,1999,16(4):444-448.
[13]周建,杨立峰,郝峰鸽,等.低温胁迫对广玉兰幼苗光合及叶绿素荧光特性的影响[J].西北植物学报,2009,29(1):136-142.
[14]张川红,沈应柏,尹伟伦,等.盐胁迫对几种苗木生长及光合作用的影响[J].林业科学,2002,38(2):27-31.
[15]周建,刘弘,尤扬,等.碱胁迫对紫荆幼苗生长与光合作用的影响[J].东北林业大学学报,2008,36(6):13-15.
[16]朱新广,张其德.NaCl对光合作用影响的研究进展[J].植物学通报,1999,16(4):332-338.
[17]吴成龙,尹金来,徐阳春,等.碱胁迫对菊芋幼苗生长及其光合作用和抗氧化作用的影响[J].西北植物学报,2006,26(3):0447-0454.
[18]崔强,王庆成,刘强,等.NaHCO3胁迫对不同胁迫时期金银忍冬苗木光合特性和荧光参数的影响[J].东北林业大学学报,2010,38(6):31-34.
[19]冯建灿,胡秀丽,毛训甲.叶绿素荧光动力学在研究植物逆境生理中的应用[J].经济林研究,2002,20(4):14-18.
[20]秦红艳,沈育杰,艾军,等.盐胁迫对不同葡萄品种叶片中叶绿素荧光参数的影响[J].中外葡萄与葡萄酒,2010(5):35-38.
[21]葛江丽,石雷,谷卫彬,等.盐胁迫条件下甜高粱幼苗的光合特性及光系统Ⅱ功能调节[J].作物学报,2007,33(8):1272-1278.
[22]张其德.盐胁迫对植物及其光合作用的影响(上)[J].植物杂志,1999(6):32-33.
