连作苎麻的部分生理生态特征及细胞学观察
2012-06-08朱四元刘头明汤清明唐守伟
朱四元,刘头明,汤清明,唐守伟
(中国农业科学院 麻类研究所,湖南 长沙 410205)
连作障碍在大豆[1]、水稻[2]、玉米[3]、小麦[4-5]、高粱[6]等多种作物栽培中普遍存在。作物连作可导致土壤速效养分含量下降、理化性状变差、土壤酶活性降低、微生物种群发生变化、产量下降等[7-11]。苎麻在生长发育过程中经常会受到连作障碍和高温干旱等不良环境的胁迫,这些因子可引起植物体内一系列生理代谢反应和生长的可逆性抑制,严重时引起植株死亡[12]。近年来,麻类的种植面积和总产量大幅度降低,其中一个主要原因是由于多年连作导致苎麻减产,麻农种麻的积极性降低[13]。笔者以中苎一号为材料,研究不同连作年限土壤种植苎麻的部分生理生态变化特征,揭示连作苎麻发生障碍效应的起始时间和表现,探索苎麻连作障碍的机理,为实现苎麻种植的可持续发展提供参考依据。
1 材料与方法
1.1 材 料
供试苎麻为中苎一号,由中国农业科学院麻类研究所提供。供试土壤为连作4~5年和8~10年的老麻园土以及从未种过苎麻的土壤,均为河潮冲积土,均取自湖南沅江试验站。
1.2 试验设计
试验于2010年1月至2011年12月在中国农业科学院麻类研究所进行。采用盆栽试验,设3个处理:连作8~10年老麻园土种植苎麻(A处理);连作4~5年土壤种植苎麻(B处理);从未种过苎麻的土壤种植苎麻(对照)。盆栽钵直径40 cm,高35 cm,每钵装土25 kg。于2010年4月中旬选取生长一致的扦插苗移栽到盆钵,每盆栽2株,每个处理30盆。在试验前期测定土壤肥力,根据测定的肥力水平对不同连作年限土壤进行后期施肥处理,使其肥力基本一致。A处理每千克土壤施20 g有机肥、0.2 g 氮肥、0.15 g磷肥、0.15 g钾肥;B处理每千克土壤施30 g有机肥、0.15 g 氮肥、0.15 g磷肥、0.15 g钾肥;C处理每千克土壤施35 g 有机肥、0.10 g氮肥、0.15 g磷肥、0.15 g钾肥。有机肥、磷肥和40%的氮肥作基肥施,与土壤充分拌匀后装入盆钵中;60%的氮肥作追肥,钾肥在苎麻旺长期一次施入。氮肥为尿素(含N 46%),磷肥为过磷酸钙(含P 12%),钾肥为氯化钾(含K 60%)。在干旱季节增加浇水次数,定期翻耕并除草。
1.3 测定项目与方法
1.3.1 土壤理化性状的测定
取不同连作年限的土壤,自然风干后过0.28 cm孔径筛。土壤有机质、全氮、全磷、有效磷、全钾、速效钾含量均采用文献[14]中的方法进行测定。
1.3.2 植株形态调查
于苎麻栽植后第1年、第2年的头麻、二麻、三麻成熟期,分别选15株测定株高、茎粗,并挖取根部,将泥土洗净烘干,测定地上植株和地下根茎的干重。
1.3.3 SOD活性和MDA含量的测定
取2011年头麻、二麻、三麻不同生育时期的叶片测定SOD活性和MDA含量[15–16]。
1.3.4 连作苎麻的显微切片观察
于2011年三麻出现连作障碍时取A处理和对照的茎杆和根部进行显微切片观察。切取苎麻茎杆(从下至上约2/3处)和根部(地下萝卜根约2/3处)长0.46 cm、宽0.35 cm的块片,采用FAA固定液(70%乙醇90.0 mL,冰醋酸5.0 mL,中性甲醛5.0 mL)固定,按文献[17]中的方法进行乙醇脱水,二甲苯透明,石蜡包埋连续切片,切片厚度为8 μm,按改良的洋红染色法[18–19]对切片进行染色,在OLYMPUS–2显微镜下观察、拍照。
1.4 数据处理
采用Excel 2003进行数据处理;采用SPSS进行差异显著性分析。
2 结果与分析
2.1 各处理土壤的理化性状
从表1可知,不同连作年限土壤的全N、全K、有效K含量以连作8~10年的最高,对照最低,有效N含量以连作8~10年的最低,对照最高。不同连作年限土壤全N、全P、有效P、有效K 含量差异不显著,连作 8~10年土壤的有机质含量显著高于连作4~5年土壤及对照,连作8~10年、4~5年的土壤有效N含量显著低于对照。

表1 不同连作年限土壤的理化性状Table 1 Changes in soil nutrients of ramie field soil under different years of continuous cropping
2.2 各处理苎麻的农艺性状
从表2可知,2010年,各处理苎麻不同时期的株高、茎粗、地上部及地下部干重、根冠比的差异不显著,未表现出连作障碍。2011年,各处理头麻的株高、茎粗、地上部干重差异不显著,A处理头麻地下部干重显著低于其他处理;A处理二麻株高显著低于B处理和对照,茎粗、地上部干重均极显著低于B处理和对照。A处理二麻地下根系基本腐烂,无法收获苎麻纤维,存在明显的连作障碍;A处理三麻收获期的连作障碍更加严重,麻株长到约40 cm时生长基本停滞。

表2 各处理苎麻的农艺性状Table 2 Agronomic traits of different treatment on ramie
2.3 各处理苎麻的SOD活性及MDA含量
从图1可以看出,2011年各处理苎麻植株SOD活性在头麻、二麻不同生育时期的变化趋势基本一致,随着植株的成熟,各处理 SOD活性均有所增加;三麻不同时期各处理间的差异比较大,成熟期A处理苎麻SOD活性显著低于B处理及对照,这与2011年A处理苎麻三麻株高、茎粗出现明显下降的连作障碍现象时期基本一致。苎麻在连作逆境条件下随着逆境时间的延长,其 SOD活性明显降低,SOD活性的变化可能导致重茬苎麻植株体内抗氧化系统遭到破坏,进而影响苎麻植株正常的生长发育。
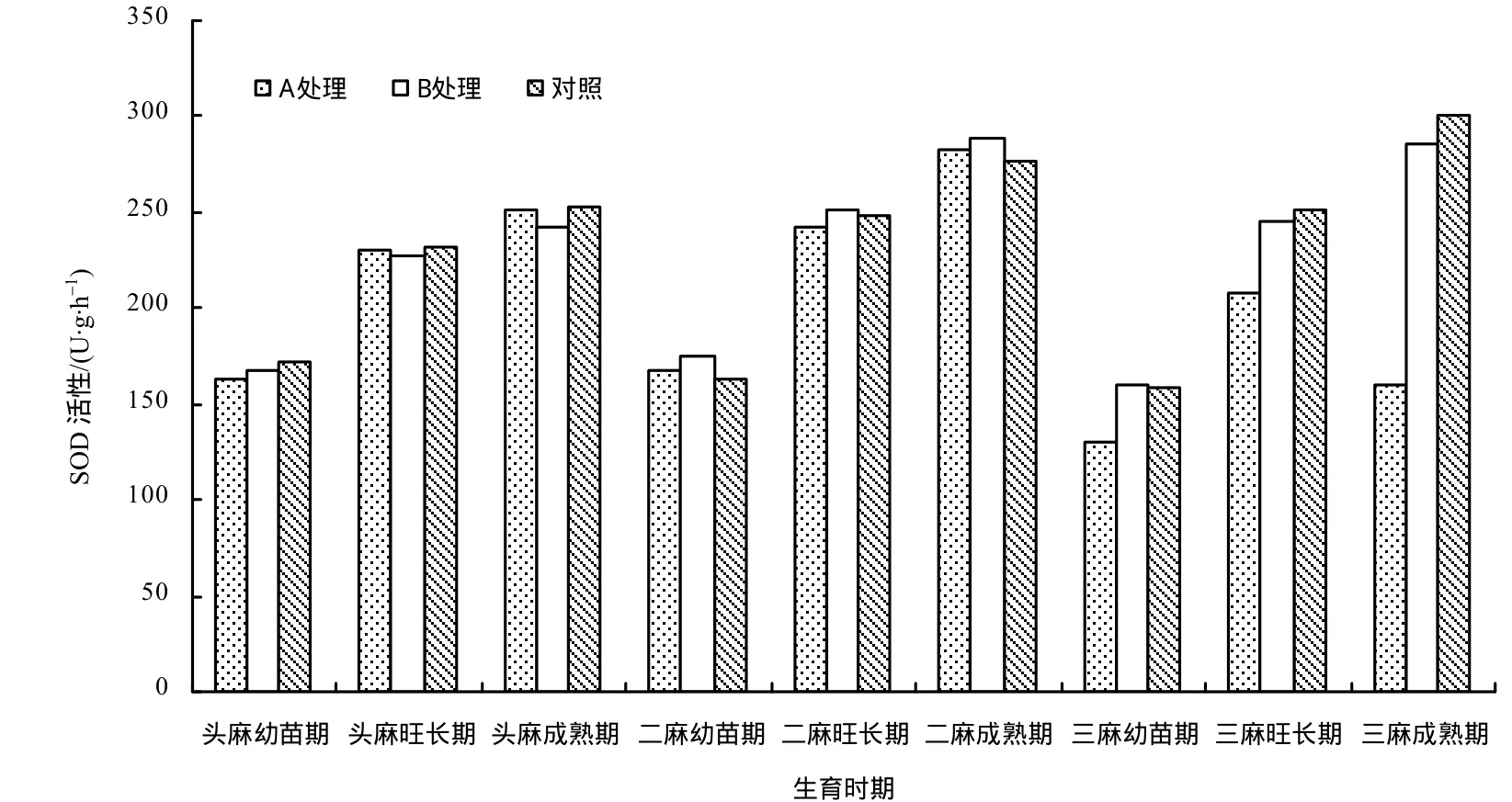
图1 各处理苎麻不同生育时期植株的SOD活性Fig.1 SOD activities of ramie at different growing of different treatments
从图2可以看出,2011年各处理苎麻植株MDA含量在头麻、二麻的不同生育时期的变化趋势基本一致,随着植株的不断成熟,MDA含量有所增加;三麻不同时期各处理间的差异比较大,在幼苗期和旺长期,A处理苎麻MDA含量高于B处理及对照,成熟期A处理显著高于B处理及对照。在连作逆境条件下,苎麻植株的MDA含量增加,苎麻叶片中产生的活性氧毒害加重,这可能是导致连作苎麻生长缓慢的原因之一。

图2 各处理苎麻不同生育时期植株的MDA含量Fig.2 MDA content of ramie at different growing of different treatments
2.4 A处理与对照苎麻显微结构的对比
从图 3–a、b可以看出,连作苎麻根部皮层薄壁细胞排列不整齐,视野中存在细胞缺失,结构分层不明显。
从图3–c、d可以看出,从外皮层往里看,A处理苎麻根部结构分层不如对照明显,连作苎麻根皮层薄壁细胞排列由致密变为疏松,视野内有细胞缺失,次生韧皮部和次生木质部之间由长方形排列整齐的薄壁细胞组成的维管形成层不够发达,维管形成层产生的新细胞数量减少,不利于麻根次生木质部和次生韧皮部的发育,导致根系吸收水分和养分的能力降低,生长缓慢,发育不良,而对照各部分结构都很明显,根系发育正常,这与盆栽试验中肉眼观察到的连作苎麻根系不如对照发达的特征相符。

图3 A处理与对照苎麻根部的显微结构Fig.3 Microstructure of the root for ramie on A treatment and the control
图4为A处理与对照苎麻茎杆的显微切片图。与对照相比,A处理苎麻皮层薄壁细胞较疏松、中柱组成部分的维管束椭圆形不明显,髓细胞体积变小,髓与髓射线不够发达,这些将影响茎杆贮藏淀粉的能力,影响植株生长,这与从外形观察到的连作植株矮小、生长缓慢的表现一致。

图4 A处理与对照苎麻茎的显微结构Fig.4 Microstructure of the stem for ramie on A treatment and the control
3 结论与讨论
本研究结果表明,不同连作年限土壤种植苎麻表现出连作障碍的程度不同。连作8~10年苎麻土壤新栽麻第2年的二麻株高显著低于对照,茎粗、地上部干重极显著低于对照,表现出明显的连作障碍现象,三麻生长基本停滞,无法收麻。连作4~5年土壤新栽麻连作障碍现象不明显。对于长期种植苎麻的地区来说,种植年限太长势必产生严重的连作障碍现象。显微切片观察发现,连作苎麻根皮层薄壁细胞排列疏松、细胞缺失、形成层结构不发达,导致苎麻地下根系储存养分和运输水分和养分的能力降低,地上部的生长受到一定抑制,连作引起根部腐烂导致植株正常生长的“源-库”关系遭到破坏是苎麻连作障碍因子之一。苎麻根系分泌他感物质、根际微生物变化等因素对苎麻连作的影响还有待进一步研究。
植物通过体内保护酶系统清除或减少逆境胁迫所产生的活性氧,避免其对组织细胞的伤害,从而表现出氧化胁迫的抗性[20]。本研究中连作苎麻SOD活性在种植第2年的二麻开始出现下降,之后在三麻幼苗期开始急剧下降,可能是因为在连作逆境下苎麻植株启动了其防御机制,使得体内SOD活性降低。一般认为,MDA在植物体内的积累是活性氧毒害的表现,其含量的多少是判断膜脂过氧化程度的一个重要指标,连作苎麻植株体内MDA含量呈上升趋势,且在3麻的旺长期显著升高,表明此时期连作苎麻叶片中产生大量活性氧和自由基,引发膜质过氧化作用,使膜的正常结构和功能受到损伤,从而导致植株生长缓慢,甚至停止生长。
[1]胡江春,王书锦.大豆连作障碍研究I.大豆连作紫青霉菌的毒素作用研究[J].应用生态学报,1996,7(4):396-400.
[2]Anaya A L,Calera M R,Mata R.Allelopathic potential of compounds isolated from Ipomoea tricolor CAV[J].Journal of Chemical Ecology,1990,16(7):2145-2152.
[3]Turco L E,Bischoff M,Breakwell D P.Contribution of soil-borne bacteria to the rotation effect in corn[J].Plant and Soil,1990,122:115-120.
[4]Guenzi W D,McCalla T M.Phenolic acids in oats,wheat,sorghum and corn residues and their phytotoxicity[J]. Agronomy Journal,1966,58(3):303-304.
[5]Lodhi M A K,Bilal R,Malik K A.Allelopathy in agroecosystems:Wheat phytotoxicity and its possible roles in crop rotation[J].Journal of Chemical Ecology,1987,13(8):1881-1891.
[6]Guenzi W D,McCalla T M.Inhibition of germination and seedling development by crop residues[J].Soil Sci Soc Am Proc,1962,26:456-458.
[7]马春梅,刘侃,唐远征,等.作物定位轮作体系长期试验研究Ⅲ.轮作与连作对土壤微生物数量的影响[J].东北农业大学学报,2005,36(2):147-152.
[8]阮维斌,王敬国,张福锁.连作障碍因素对大豆养分吸收和固氮作用的影响[J].生态学报,2003,23(1):22-29.
[9]孙秀山,封海胜,万书波,等.连作花生田主要微生物类群与土壤酶活性变化及其交互作用[J].作物学报,2001,27(5):617-621.
[10]于广武,许艳丽,刘晓冰,等.大豆连作障碍机制研究初报[J].大豆科学,1993,3:24-29.
[11]张淑香,高子勤,刘海玲.连作障碍与根际微生态研究Ⅲ.土壤酚酸类及其生物学效应[J].应用生态学报,2000,11(5):741-744.
[12]Liu Fei-hu,Liu Qi-yuan,Liang Xue-ni,et al. Morphological,anatomical,and physiological assessment of ramie [Boehmeria Nivea (L.)Gaud.]tolerance to soil drought[J].Genetic Resources and Crop Evolution,2005,52:497–506.
[13]毛长文,毛宗礼.湖北省苎麻生态产业发展面临的问题与对策[J].安徽农业科学,2010,38(24):13469–13470,13512.
[14]鲍士旦.土壤农化分析[M].第3版.北京:农业出版社,2005.
[15]刘祖琪,张石城.植物抗性生理学[M].北京:中国农业出版社,1992:371.
[16]Heath R L,Packer L.Photoperoxidation in isolated chloroplasts I.Kinetics and stoichiometry of fatty acidperoxidation[J].Archives of Biochemistry and Biophysics,1968:125,189-198.
[17]郑国锠.生物显微技术[M].北京:人民教育出版社,1979.
[18]Zhou Z H,Yu P,Liu G H,et al.Morphological and molecular characterization of two G.somalense monosomic alien addition lines(MAALs)[J].Chinese Science Bulletin,2004,49(9):910-914.
[19]周仲华,何鉴星,陈金湘.陆×索棉异附加单体系的形态学及细胞学鉴定[J].湖南农业大学学报:自然科学版,2004,30(4):316-318.
[20]Lara L,Luca S.Copper toxicity in Prunus cerasifera:Growth and antioxidant enzymes responses of in vitro grown plants[J].Plant Science,2005:168,797–802.
