六盘山主要树种叶片稳定性碳同位素组成的时空变化特征
2012-05-07王云霓王彦辉程积民于澎涛徐丽宏李振华张晓蓓
王云霓,熊 伟,王彦辉,程积民,于澎涛,徐丽宏,何 聪,郝 佳,李振华,张晓蓓
(1.中国林业科学研究院森林生态环境与保护研究所,北京100091;2.中国科学院水利部水土保持研究所,陕西 杨凌712100;3.广西大学 林学院,南宁530005;4.河北农业大学 林学院,河北 保定071000)
植物体中稳定性碳同位素组成(δ13C)是反映植物胞间与大气CO2之间分压比(Ci/Ca)的一个整合指标[1]。由于植物在叶片的光合过程中,会产生碳同位素分馏现象,从而叶片更趋向于同化12CO2而排斥13CO2,这使得叶片中的碳同位素丰度远低于大气CO2。因而,δ13C值常被用来表征植物叶片的光合能力及水分利用效率(water use efficiency,简称WUE)。
在自然界中,植物的δ13C值与其生物学特性和环境因子密切相关[2-4]。植物首先会通过遗传特性或代谢特征来影响其δ13C值的大小[2,5]。如δ13C值的种间[6]和生活型差异[7-8]就反映了其生物学特性之间的区别。同时,植物还会受环境因子所影响的叶片净光合速率和气孔导度导致其δ13C的变化[1]。例如,同一种植物在不同水分生境的个体[9-11]和同一个体在不同冠层部位或方位[12-16]之间的δ13C值差异则说明了环境因子的影响。另外,受季节变化所导致的辐射、降水和温度等环境因子的变化和植物物候节律的双重影响,也会导致植物叶片δ13C值的变化。严昌荣等[3]研究表明北京山区六种木本植物的δ13C和WUE在早春时高于生长旺期;而王玉涛等[8]发现落叶阔叶乔木叶片的δ13C值表现出春季>秋季>夏季的规律,但常绿针叶乔木叶片的δ13C值变化不大。综上所述,尽管目前尽管对叶片δ13C值的种内、种间及季节变化进行了广泛的研究,但因各研究的对象、地点、采样时间不一致,其结论还存在着一定的分歧。鉴于以上认识,可以认为六盘山区内主要树种叶片δ13C值是存在种间、种内差异,但这种差异会随着其季节变化特征及水分环境条件的改变而变化的。
宁夏六盘山林区位于黄土高原中西部,地处半湿润半干旱地区的过渡带,显著的水源涵养效益使之被誉为该区的“湿岛”,对宁南山区气候调解和生态平衡保护起着十分重要的作用[17]。过去,由于对该区森林植被的严重破坏,导致了生态环境的逐步恶化,所以急需选择节水树种、合理配置结构来恢复当地的森林植被及其生态功能[18-20],而在干旱半干旱地区水源涵养林的营建中要尽可能的选择低耗水、高WUE的树种。目前,在六盘山林区针对典型植被和主要树种的水分生理参数[21]、树干液流[22]和蒸散[23]等方面做了一些工作,但还没有该区主要树种叶片的δ13C值及WUE方面的研究。
本文以六盘山叠叠沟小流域(代表半干旱区)和香水河小流域(代表半湿润区)为研究区域,以该区的主要树种为对象,在生长季的前期、中期和后期分别测定主要树种叶片的δ13C,主要目的在于探讨:(1)同一树种叶片δ13C的冠层部位和方位差异;(2)不同树种叶片δ13C的差异及季节变化;(3)同一树种不同水分生境下叶片δ13C的差异。
1 材料与方法
1.1 研究区概况
试验样地设在六盘山南侧的香水河小流域和北侧的叠叠沟小流域。香水河小流域 (106°09′—106°30′E,35°15′—35°41′N)位于六盘山南侧,海拔2 040~2 931m;属半湿润气候,热量较低,水分充足,近10a的年均气温为6.8℃,年均降水量636.8mm,多集中于7—9月(图1)。主要树种有华山松(Pinus armandii)、辽东栎 (Quercus liaotungensis)、白桦(Betula platyphylla)、少脉椴(Tilia paucicostata)、油松(Pinus tablaeformis)、红桦(Betula albo-sinensis)、华 北 落 叶 松 (Larix principris-rupprechtii)、山 桃(Prunus davidiana)和沙棘(Hippophae rhamnoides)等。叠叠沟小流域(106°04′—106°09′E,35°54′—35°58′N)位于六盘山北侧,海拔1 975~2 615m;属半干旱气候,近10a的年均气温为7.8℃,年均降水量413.5 mm,主要集中在7—9月(见图1),主要树种有华北落叶松、沙棘和山桃等。2个小流域的土壤类型均以山地灰褐土为主,土层厚度为30—100cm。
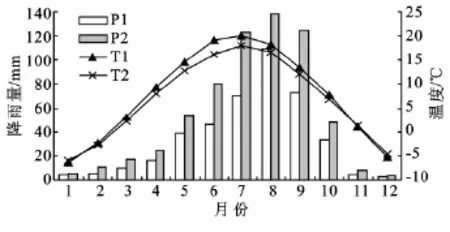
图1 六盘山近10a(2001-2010年)的月平均降水量和温度分布(P1、T1:半干旱区降水量和气温;P2、T2:半湿润区降水量和气温)
1.2 试验设计与样品采集
为分析不同树种叶片的δ13C差异及季节变化,于2010年6月17—18日(生长季前期)、7月30—8月5日(生长季中期)和9月9—13日(生长季后期),在香水河小流域选择辽东栎、白桦、少脉椴、华山松、油松、华北落叶松、沙棘、山桃等8个主要树种(见表1),每个树种各选取4株长势良好的树木为样木,每株样木选取树冠南向中上部枝条的叶片取样,样品为4株样木叶片的混合样。
为分析同一树种叶片δ13C的冠层部位差异,在生长季中期,采集辽东栎、白桦、华山松、油松、华北落叶松5个树种冠层的上部、中部和下部南向的叶片样品;同时,在这5个树种冠层中部的东、西、南、北等4个方位分别取样,用以比较同一树种冠层不同方位叶片δ13C的差异。为分析不同水分生境下树木叶片δ13C的种内个体差异及季节变化,于生长季的初、中、末期在叠叠沟小流域和香水河小流域分别选取华北落叶松和沙棘2个树种的树冠南向中上部枝条的叶片分别采样。
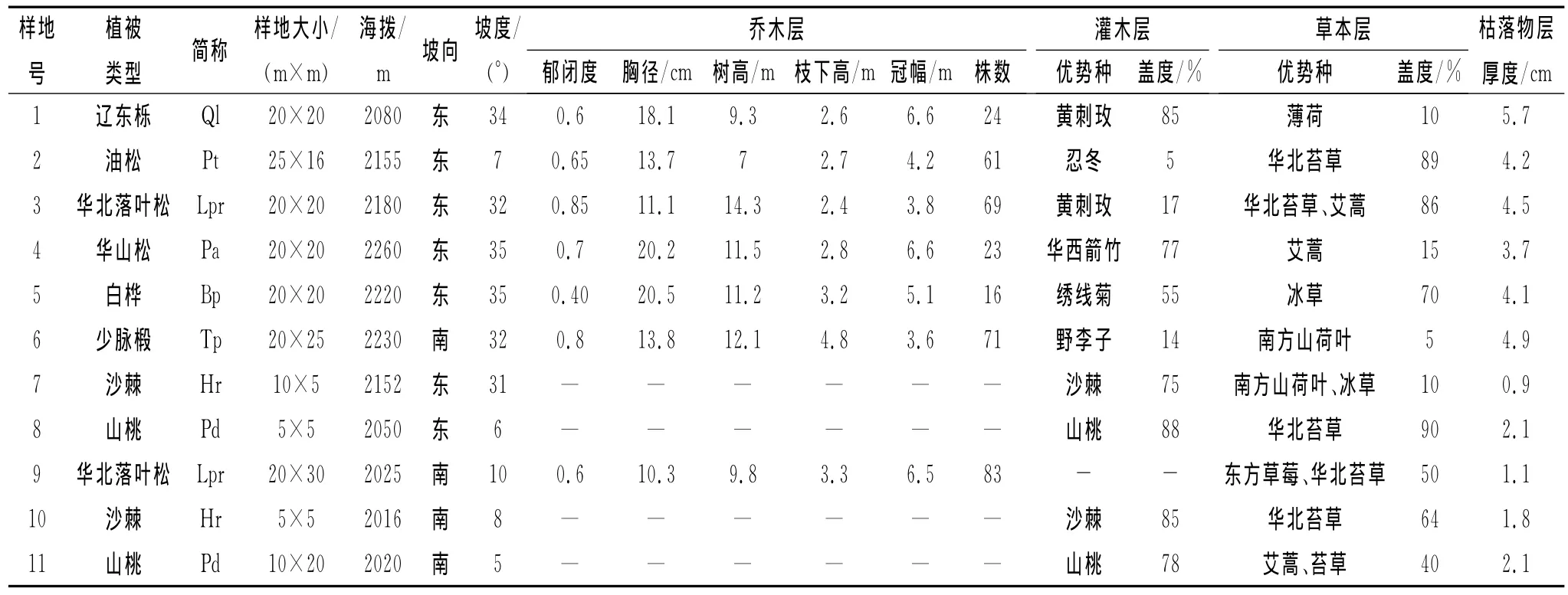
表1 六盘山香水河小流域和叠叠沟小流域主要树种所在样地的植被基本情况
1.3 样品处理与分析
将采回叶片样品置于恒温干燥箱中,105℃杀青,60℃烘干(连续48h),粉碎后过80目筛制成样品,然后进行样品的稳定碳同位素比值分析(中国林业科学研究院稳定同位素比率质谱实验室,精度为0.1‰)。通过DELTA V Advantage同位素比率质谱仪和Flash EA1112HT元素分析仪(Thermo Fisher Scientific,Inc.,USA)测定CO2的13C与12C比率,并与国际标准物PDB(Pee Dee Belemnite)比对后计算样品的δ13C值,根据Farquhar等[1]的研究,植物的δ13C值可由下式来计算:

式中,(13C/12C)sample和(13C/12C)PDB分别为测试样品和标样PDB的δ13C值。
1.4 数据处理
用SPSS 16和Excel 2003对数据进行统计分析。
2 结果与分析
2.1 主要树种冠层不同部位叶片δ13C的差异分析
图2表明,各树种不同冠层部位的叶片δ13C值均存在差异,且不同树种间也存在差异。其中,华北落叶松、白桦和油松等3个树种叶片δ13C值垂直分布表现为冠层上部>中部>下部,上部叶片的δ13C值分别是-27.19‰,-27.44‰和-27.01‰,比下部高出了0.72‰,0.93‰和0.49‰;但华山松和辽东栎叶片的δ13C垂直分布表现为冠层下部>上部>中部,其中上部叶片的δ13C值分别是-26.23‰,-27.23‰,比下部高出了0.18‰,0.07‰,说明叶片δ13C值的冠层垂直变化存在着种间差异,可能与树种的生物学特性及其小生境有关。
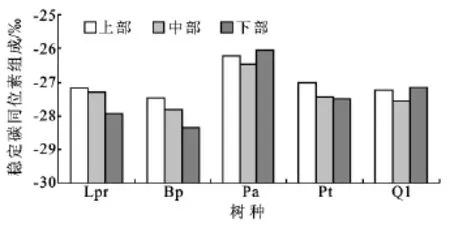
图2 六盘山香水河小流域主要树种叶片δ13C的冠层高度变化
图3表明,各个树种叶片δ13C值在不同冠层方位也存在着差异,但差异较小相对,一般在2.8%以下。具体来说,华北落叶松和白桦为南向>东向>北向>西向,油松和辽东栎为西向>南向>北向>东向,华山松为北向>东向>南向>西向,这可能与树木不同方位光照条件差异所导致的树木生理活动差异有关。

图3 六盘山香水河小流域主要树种同一冠层高度叶片δ13C的方位变化
2.2 主要树种叶片δ13C的季节变化对比分析
图4和图5显示了六盘山辽东栎、白桦、少脉椴、华山松、油松、华北落叶松、沙棘、山桃等8个主要树种叶片δ13C的季节变化。结果表明,各树种叶片的δ13C值在生长季初期较高,而在生长季中、后期较低,但其随水分生境条件变化而异。在香水河小流域,各树种叶片的δ13C值在生长季初最高,而后持续降低,到生长季后期到达最低值,但各树种叶片的δ13C值下降幅度不同(图4);在叠叠沟小流域,华北落叶松、沙棘和山桃等3种树种叶片δ13C值也是生长季初期最高,但在生长季中期树种叶片的δ13C值最低,后期略有回升,表现为“高—低—高”的变化特征(图5)。

图4 六盘山香水河小流域主要树种叶片δ13C季节变化

图5 六盘山叠叠沟小流域主要树种叶片δ13C季节变化
2.3 不同水分生境条件下同一树种叶片δ13 C的对比分析
以华北落叶松和沙棘为例,分析了同一树种不同个体的叶片δ13C对不同水分生境条件的响应。在不同水分生境下,两个树种叶片的δ13C存在着明显种内个体差异,即对于同一树种,生长在叠叠沟小流域树木叶片δ13C值均大于生长在香水河小流域的,说明同种树木生长在半干旱地区的叶片WUE大于生长在半湿润地区的(图6和图7)。然而,在不同水分生境下同一树种叶片δ13C值的差异大小随着季节而变化,即在生长季初、末期,δ13C差异较大,而生长季中期差异则十分微小。

图6 华北落叶松在两种水分生境条件下的叶片δ13C值对比
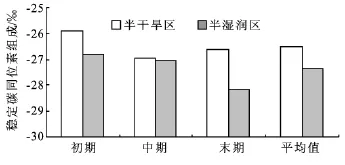
图7 沙棘在两种水分生境条件下的叶片δ13C值对比
3 讨论
3.1 主要树种叶片δ13C的冠层差异分析
大量研究表明,树木不同冠层部位叶片所接受的光照时间、强度及温湿度存在着差异[16,24],而这些微气象因子差异又可能导致叶片的气孔导度(Gs)和光合能力等发生变化,从而引起叶片δ13C值之间的差异[13-14]。一般地,树木冠层上部的光照条件较好,其光合速率相对较高,光合同化的CO2增多,而Gs相对较低,使叶片内部的CO2浓度相对较低,从而造成叶片δ13C值相对较高。如Duursma&Marshall[13]测定了美国北部落基山脉混合林中9种针叶树种的200多株样树的叶片δ13C,发现其从树冠的上部到下部逐渐减小,并指出光合能力变化是引起δ13C垂直变化的决定因素。Koch等[12]发现北美红杉(Sequoia sempervirens)的叶片δ13C从树冠40m上升到110m时增大了5‰。本研究中,华北落叶松、白桦和油松等3个树种叶片δ13C值的冠层变化表现为上部>中部>下部,这与前人研究的结果一致[13-14,24],说明上述3个树种的光照条件在冠层部位的垂直变化导致了其叶片δ13C的差异。另外,同一树种叶片δ13C的冠层差异可能还与树木的水力结构分布[25]、叶片形态结构[26]和土壤呼吸[27]的变化有关,但这还有待于进一步研究。
本研究中,华山松和辽东栎冠层下部的δ13C值却大于冠层中、上部,这可能是因为华山松和辽东栎的光饱和点较低,7月份下旬研究区域的光强和温度较高,细胞内外蒸气压亏缺增大的影响,使其冠层上、中部叶片内光合羧化酶的活性受到抑制,从而消弱了树木的光合作用;而冠层中下部的光强相对较弱,光合羧化酶的活性受到抑制程度较低,光合速率相对较大,同时由于在高光强条件下,中、下层叶片的蒸腾速率明显小于上层叶片[28],故其表现为较高的δ13C值和WUE。
3.2 不同水分生境条件下同一树种叶片δ13C的对比
植物对不同水分环境具有一定的弹性适应能力[29]。一般地,在水分条件较差的生境中,植物经过一定的抗旱锻炼,会通过形成与环境相适应的形态结构和生理生化机制,来提高其WUE,从而增强对干旱的适应能力[30-33]。文陇英等[11]测定了不同水分生境下祁连圆柏(Sabina przewalskii)叶片的δ13C值,发现在相对干旱的环境下δ13C值高于相对湿润环境,并指出半干旱生境中的植物会通过改变脯氨酸、叶绿素等含量来提高植物的WUE,增强其抗旱能力。陈世萍等[9]测定了内蒙古锡林河流域6个不同水分条件下的典型植物群落中黄囊苔草(Carex korshinskyi)叶片δ13C值、含水量及其种群特征变化,结果表明,随着土壤水分的降低,叶片δ13C值显著增大,且不同生境下的黄囊苔草能够通过改变其WUE来适应不同的土壤水分状况,使其植物群落中表现出更强的竞争能力和生态适应性。Peuke等[10]比较了来自不同气候条件下山毛榉(Fagus sylvatica)幼苗的δ13C值,发现干旱胁迫显著增加了叶片的δ13C值。本研究与以上研究的结论基本一致,即生长在半干旱生境下的华北落叶松和沙棘叶片δ13C值均显著大于半湿润生境(P<0.05),说明这两树种在半干旱区表现出更高的WUE,这种生存策略可能是植物对不同水分生境差异长期适应的结果。然而,本研究还发现在不同水分生境下,同一树种叶片δ13C值的种内个体差异会随着季节而变化,即在生长季初、末期,同一树种的这种δ13C差异较大,而生长季中期这种差异十分微弱。这是由于研究区降雨多集中在生长季中期,使两个小流域内的植物都能获得较多的可利用水分,从而使叶片δ13C值较低且差异不明显。Sun等[34]对生长在南京、北京和兰州这三个地区银杏(Ginkgo biloba L.)的δ13C值进行了研究,结果表明,随降水量减少叶片δ13C呈逐渐增加趋势;而Stewart等[35]对澳大利亚东部、陈拓[36]对阜康和金塔荒漠、Zheng等[37]对黄土高原的C3植物δ13C随降水量变化的研究也都表明,降雨量的增加会使叶片δ13C值和WUE减小。以上事实说明,不同生境区域内降雨量的季节分配会对同一种植物叶片的δ13C值和WUE产生较大的影响。
4 结论
(1)乔木树种叶片的δ13C值存在冠层差异,但各树种的冠层垂直变化特征不尽相同。本研究中,华北落叶松、白桦和油松等3个树种叶片δ13C值表现为上部>中部>下部;而华山松和辽东栎为冠层下部>上部>中部,但各树种叶片的δ13C方位差异不显著。这说明在今后的对于树木叶片δ13C的研究中,应该更加注重其随树高的垂直变化所带来的差异。
(2)六盘山各主要树种叶片δ13C值都存在季节变化,在生长初期各树种叶片δ13C值较高,而在生长季中、后期较低,其变化也随水分生境条件不同而异,在半湿润区叶片δ13C在整个生长季持续降低,在半干旱区却表现为“高—低—高”的变化趋势。
(3)在不同水分生境条件下,华北落叶松和沙棘叶片的δ13C值存在着明显种内个体差异,但这种差异受降雨量的季节分配影响较大。旱季(生长季初、末期),同一树种在半干旱区表现出更高的δ13C,而在雨季(生长季中期)δ13C相接近。因此,在造林实践中,可对树木幼苗采取适度的抗旱锻炼来提高其对干旱环境的适应能力。
[1] Farquhar G D,O′Leary M H,Berry J A.On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves[J].Australian Journal of Plant Physiology,1982,9(2):121-137.
[2] Button T W,Shin-ichi Yamsaki.Stable carbon isotope ratios of soil organic matter and their use as indicators of Vegetation and Climate Change[M].Marcel Dekker,Inc,New York,1996:47-82.
[3] 严昌荣,韩兴国,陈灵芝.六种木本植物水分利用效率和其小生境环境关系研究[J].生态学报,2001,21(11):1952-1956.
[4] 冯元虎,安黎哲,陈拓,等.马先蒿属(Pedicularis L.)植物稳定碳同位素组成与环境因子的关系[J].冰川冻土,2003,25(1):88-93.
[5] 韩兴国,严昌荣,陈灵芝,等.暖温带地区几种木本植物碳稳定同位素的特点[J].应用生态学报,2000,11(4):497-500.
[6] Chevillat V S,Siegwolf R T W,Pepin S,et al.Tissuespecific variation ofδ13C in mature canopy trees in a temperate forest in central Europe[J].Basic and Applied E-cology,2005,6(6):519-534.
[7] Chen Shiping,Bai Yongfei,Han Xingguo.Variations in composition and water use efficiency of plant functional groups based on their water ecological groups in the Xilin river basin[J].Acta Botanica Sinica,2003,45(10):1251-1260.
[8] 王玉涛,李吉跃,刘平,等.不同生活型绿化植物叶片碳同位素组成的季节特征[J].植物生态学报,2010,34(2):151-159.
[9] 陈世苹,白永飞,韩兴国,等.沿土壤水分梯度黄囊苔草碳同位素组成及其适应策略的变化[J].植物生态学报,2004,28(4):515-522.
[10] Peuke A D,Gessler A,Rennenberg H.The effect of drought on C and N stable isotopes in different fractions of leaves,stems and roots of sensitive and tolerant beech ecotypes[J].Plant,Cell & Environment,2006,29(5):823-835.
[11] 文陇英,陈拓,张满效,等.不同生境下祁连圆柏叶片色素和稳定碳同位素组成的变化[J].冰川冻土,2010,32(4):823-828.
[12] Koch G W,Sillett S C,Jennings G M,et al.The limits to tree height[J].Nature,2004,428:851-854.
[13] Duursma R A,Marshall J D.Vertical canopy gradients in δ13C correspond with leaf nitrogen content in a mixed-species conifer forest[J].Trees-Struoture and Function,2006,20(4):496-506.
[14] Woodruff D R,Mcculloh K A,Warren J M,et al.Impacts of tree height on leaf hydraulic architecture and stomata control in Douglas-fir[J].Plant,Cell & Environment,2007,30(5):559-569.
[15] Ayumi Tanaka-Oda,Tranaka Kenzo,Satomi Koretsune,et al.Ontogenetic changes in water-use efficiency(δ13C)and leaf traits differ among tree species growing in a semiarid region of the Loess Plateau,China[J].Forest Ecology and Manage,2010,259(5):953-957.
[16] 何春霞,李吉跃,张燕香,等.5种绿化树种叶片比叶重、光合色素含量和δ13C的开度与方位差异[J].植物生态学报,2010,28(7):3008-3016.
[17] 张智,林莉,孙银川,等.全球变暖对宁夏六盘山区气候的影响[J].干旱区资源与环境,2007,21(8):64-67.
[18] 程积民,万惠娥.中国黄土高原植被建设与水土保持[M].北京:中国林业出版社,2002.
[19] 王彦辉,金旻,于澎涛.我国与森林植被和水资源有关的环境问题及研究趋势[J].林业科学研究,2003,16(6):739-747.
[20] 张文辉,刘国彬.黄土高原地区植被生态修复策略与对策[J].中国水土保持科学,2009,7(3):114-118.
[21] 刘海龙,余治家,熊伟,等.宁夏六盘山10种木本植物抗旱性研究[J].中国水土保持科学,2009,7(1):62-66.
[22] 熊伟.六盘山北侧主要造林树种水分消耗特征研究[D].北京:中国林业科学研究院,2003.
[23] Wang Yanhui,Yu Pengtao,Xiong Wei.Water-Yield reduction after afforestation and related processes in the semiarid Liupan Mountains,Northwest China[J].Journal of the American Water Resources Association,2008,44(5):1086-1097.
[24] Powers M D,Pregitzer K S,Palik B J,et al.Water relation of pine seedlings in contrasting overstory environments[J].Forest Ecology and Management,2009,258(7):1442-1448.
[25] Niinemets U,Tenhunen J D,Beyschlag W.Spatial and age-dependent modifications of photosynthetic capacity in four Mediterranean oak species[J].Functional Plant Biology,2004,31(12):1179-1193.
[26] 何春霞,李吉跃,郭明,等.4种乔木叶片光和特性和水分利用效率随树高的变化[J].生态学报,2008,28(7):3008-3016.
[27] Brooks J R,Flanagan L B,Vareny G T,et al.Vertical gradients in photosynthetic gas exchange characteristics and reification of respired CO2within boreal forest canopies[J].Tree Physiol,1997,17(1):1-12
[28] 肖文发,徐德应,刘世荣,等.杉木人工林针叶光合与蒸腾作用的时空特征[J].林业科学,2002,38(5):38-46.
[29] 张淑勇,周泽福,张光灿,等.水分胁迫下天然次生灌木山桃和山杏光合气体交换特征[J].西北植物学报,2008,28(12):2492-2499.
[30] 孙志虎,王庆成.土壤含水量对三种阔叶树苗气体交换及生物量分配的影响[J].应用与环境生物学报,2004,10(1):7-11.
[31] 张岁岐,周小平,慕自新,等.不同灌溉制度对玉米根系生长及水分利用效率的影响[J].农业工程学报,2009,25(10):1-6.
[32] 崔晓军,薛昌颖,杨晓光,等.不同水分处理旱稻农田蒸散特征和水分利用效率[J].农业工程学报.2008,24(4):49-54.
[33] 彭强,梁银丽,陈晨,等.辣椒叶片生理特性对光照强度和土壤水分的响应[J].农业工程学报,2010,26(增刊1):115-121.
[34] Sun B N,Dilcher D L,Beerling D J,et al.Variation in Ginkgo biloba L leaf characters across a climatic gradient in China[J].Pnas,2003,100(12):7141-7146.
[35] Stewart G R,Turnbull M H.13C natural abundance in plant communities along a rainfall gradient:a biological integrator of water availability[J].Funotional Plant Biology,1995,22(1):51-55.
[36] 陈拓,冯虎元,徐世建,等.荒漠植物叶片碳同位素组成及其水分利用效率[J].中国沙漠,2002,22(3):288-291.
[37] Zheng Shuxia,Shangguan Zhouping.Spatial patterns of foliar stable carbon isotope compositions of C3plant species in the Loess Plateau of China[J].Ecological Research,2007,22(2):342-353.
