高寒草甸恢复演替过程中生产力与植物功能特征关系的研究
2012-04-12张灵菲魏斌葛庆征郝敏傅华张卫国江小雷
张灵菲,魏斌,葛庆征,郝敏,傅华,张卫国,江小雷
(兰州大学草地农业科技学院 草地农业生态系统国家重点实验室,甘肃 兰州730020)
*植物功能特征是指那些直接影响生态系统过程或生态系统响应环境条件变化的物种特征[1-3],如叶片的大小、叶面积;种子大小、种子扩散模式;植物高度、生长速率;根的长度、根系类型和固氮性能等。植物功能特征(plant functional traits)对生态系统功能及生态系统过程响应全球变化有重要影响[4]。不同的物种,因其功能特征的不同,对生态系统功能产生的影响会截然不同。研究表明,植物的功能特征是植物在长期进化过程中对环境变化的适应性表现,对生态系统功能有一定的指示作用[5,6]。植物的功能特征可直接影响能量流通和物质循环,间接改变非生物条件,进而调节生态系统过程[7]。越来越多的研究表明,植物功能特征是联系物种与生态系统的纽带[8,9],因此,在探讨物种多样性的变化、物种成分的变化对生态系统功能的影响时,需要更多的考虑物种的功能特征所发挥的作用[3,8]。
作为初级生产者,植物可通过其物种数量、物种丰富度、物种成分等多个方面对生态系统功能产生影响[10,11],由于这些因素具有不同的表征属性:有的是数量特征,有的是质量特征,因而在探讨植物群落与生态系统关系时,难以同时考虑不同因素的综合作用[12,13]。Garnier等[6]提出的“生物量比例理论”(biomass ratio hypothesis)则较好地解决了这一难题。生物量比例理论认为,物种的功能特征对生态系统性质的影响程度与该物种的生物量在群落中所占比例密切相关,因而可将物种数、物种丰富度以及物种成分等各种因素通过功能特征和生物量的关系联系在一起,进而实现对特定物种在生态系统中发挥的具体作用进行量化评定[9]。这意味着生态系统功能在很大程度上取决于优势物种的功能特征及其在群落中所占的生物量。
高寒草甸作为青藏高原的主要草地类型,不仅是畜牧业的资源依托,而且也是欧亚大陆极具特色的生态系统类型。但由于长期的粗放经营,超载过牧,以及对草地资源的不合理开发利用,使部分草地的原生植被严重退化,从而形成大量处于不同恢复演替(次级演替)阶段的草地。次生演替是草地生态系统中普遍存在的现象,也是研究植物功能特征响应环境变化的理想场所[6]。利用退化高寒草甸研究草地植物群落的演替对植物功能特征变化与生态系统功能关系的影响,对深刻认识草地生态系统的本质及草地的可持续利用有重要意义。本研究的主要目的是利用生物量比例理论,研究高寒草甸恢复演替过程中草地生产力与植物群落优势物种功能特征的关系,以期对生物量比例理论进行验证,同时探讨植物功能特征对高寒草甸生态系统功能响应环境变化的指示意义。
1 材料与方法
1.1 实验地概况
研究地设于甘肃省玛曲县阿孜牧场所辖区内。地理坐标为N 33°43′45″,E 101°44′50″,海拔3 500 m。土壤为亚高山草甸土。年均温1.4℃,年均降水量463.9 mm。植被类型为亚高山草甸,属冬季牧场,草地退化明显。主要植物组成为垂穗披碱草(Elymusnutans)、洽草(Koeleriacristata)、矮嵩草(Kobresiahumilis)、甘肃嵩草(K.kansuensis)、鹅绒委陵菜(Potentillaanserrina)、二裂委陵菜(P.bifurca)、马先蒿(Pedicularissp.)、乳白香青(Anaphalislactea)、细叶亚菊(Ajaniatenuifolia)、草玉梅(Anemonerivularis)等。
1.2 样方设定及植物功能特征的测定
用空间序列代替时间序列的方法,分别于同质高寒草甸内选择弃耕后恢复1,3,7,10,15年的5个演替梯度的草地各0.5 hm2,于每梯度样地内选5个0.5 m×0.5 m的样方,分别于2010年5月(牧草萌发期)和8月底(生物量高峰期)分种测定植物的高度、盖度、密度和生物量。同时按照生物量比率理论[6],测定样方中生物量之和达80%的优势物种的叶面积、叶鲜重,然后将样品带回实验室60℃条件下烘干,称各物种的干重,用以测定和计算相关数据。
研究选择4种与养分利用、生产力关系密切且简单易测的植物功能特征[9]:植株高度 (plant height,PH,植物的自然高度,cm)、相对生长率 (relative growth rate,RGR)、比叶面积 (specific leaf area,SLA,植物叶面积/叶干重,cm2/g),叶干物质含量 (leaf dry mass content,LDMC,叶鲜重/烘干重,g/g)。SLA 和 LDMC按照Garnier等[14]介绍的方法测定;相对生长率按照 Hunt[15]的方法计算:
式中,W f为最终生物量,W i为初始生物量,t为2次测定的间隔天数。
植物群落的集合功能特征用下式计算[16]:

式中,T为群落的功能特征值,Pi是种i的生物量,traiti是种i的特征值,n是优势物种数。
选择反映生态系统碳和能量输入性能的净初级生产力(net primary productivity,NPP)作为生态系统功能的替代指标,其计算公式为:
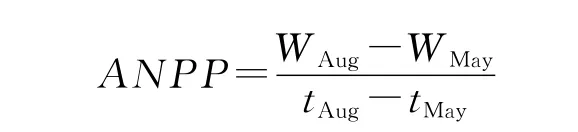
式中,ANPP为地上净初级生产力(g/m2·d),WMay为5月地上生物量(g),WAug为8月地上生物量,t为测定时间(d)。
1.3 统计分析
使用SPSS 13.0统计分析软件对数据进行统计处理,利用简单线性相关模型对演替梯度与植物功能特征的关系、演替梯度与初级生产力的关系及群落功能特征与生产力关系进行分析,利用方差分析 (one-way ANOVA)检验相关显著性。
2 结果与分析
2.1 群落功能特征与演替的关系
植物群落各功能特征对演替时间的反应有所不同。相关分析表明(图1),植株高度 (y=-12.299x+410.3,R2=0.678 2,P<0.001)、相对生长率 (y=-0.022 9x+0.624 7,R2=0.699 2,P<0.001)以及比叶面积(y=-61.171x+2 756.5,R2=0.595 6,P<0.001)等功能特征均随演替时间的延长而下降,其与演替时间呈显著的负相关关系,而叶干物质含量则随演替时间的延长而增加,二者间呈显著的正相关关系 (y=0.170 5x+0.106 3,R2=0.506 5,P<0.001)。表明在植物群落演替过程中,物种的功能特征随着物种组成的变化而发生了变化。
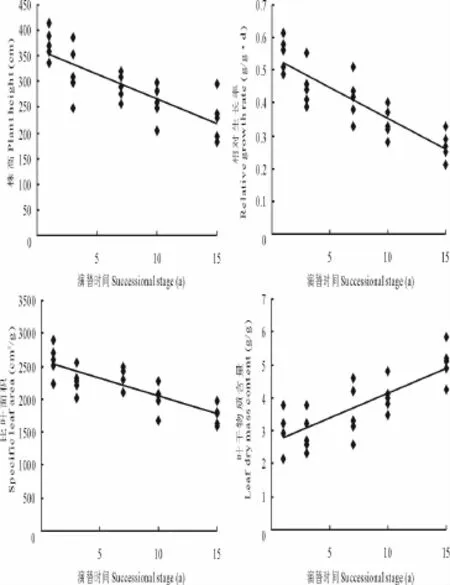
图1 群落功能特征与演替的关系Fig.1 Relationship between aggregated functional traits and succession
2.2 草地净初级生产力与演替的关系
在演替初期,地上净初级生产力水平较高,而随着演替时间的延长,地上净初级生产力呈下降趋势,二者间呈显著的负相关关系 (y=-0.228x+7.386 7,R2=0.495 4,P<0.01,图2)。表明随着演替时间的增加,草地生态系统生产功能有减弱的趋势。
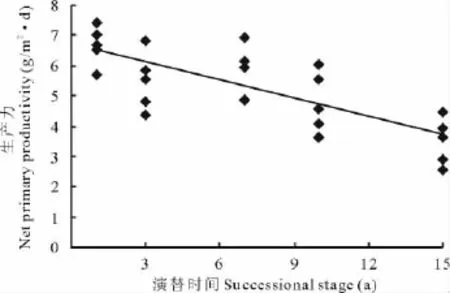
图2 草地净初级生产力与演替的关系Fig.2 Relationship between ANPP and succession
2.3 草地净初级生产力与群落功能特征的关系
地上净初级生产力随株高(y=0.014 7x+0.942 6,R2=0.457 5,P<0.01,图3)、相对生长率(y=8.695 4x+1.869 9,R2=0.519 1,P<0.001)和比叶面积 (y=0.002 4x+0.942 6,R2=0.361 7,P<0.01)的增加而增加,生产力分别与该3个功能特征呈显著正相关关系。而地上净初级生产力与叶干物质含量的关系则有所不同,随着叶干物质含量的增加,净初级生产力呈下降趋势,但二者间相关关系不显著(y=-0.435 5x+6.797,R2=0.101 5,P>0.05,图3)。表明株高、相对生长率和比叶面积等功能特征对地上净初级生产力有促进作用,而叶干物质含量与净初级生产力间没有直接关系。
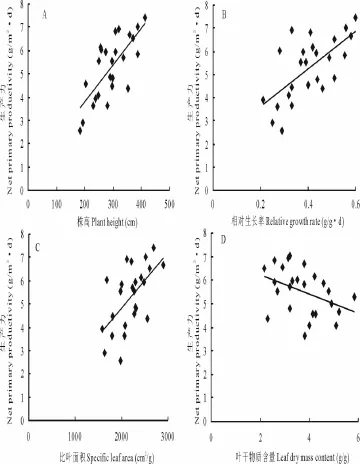
图3 地上净初级生产力与群落功能特征的关系Fig.3 Relationship between ANPP and aggregated functional traits
3 讨论
演替是草地生态系统遭受干扰后出现的普遍现象,演替草地是研究物种多样性,群落结构及各种生态过程的理想场所[17,18]。本研究利用高寒草甸这一独特草地类型,探讨草地恢复演替过程中植物群落优势物种的功能特征与地上净初级生产力的关系,结果表明,与生产力密切相关的植株高度、相对生长率和比叶面积等功能特征值均随着演替的进程而下降 (图1,3)。此结果与其他不同类型草地生态系统演替的研究类似,表明物种的功能特征随着演替时间发生变化的趋势,体现出演替早期,植物群落由获取养分能力较强、生长速度较快的物种(具有较高的相对生长率、较大的比叶面积值和较低的叶干物质含量的物种)占主导地位,而演替后期,这类物种逐渐由生长较慢、竞争力较强的物种(具有较低的相对生长率、较小的比叶面积和较高的叶干物质含量值的物种)所取代,而生态系统功能也随之减弱[19-21]。
不同的物种其功能特征值有所不同,群落集合功能特征随着演替而发生显著的变化,一定程度上表明具有不同功能特征值的物种组成相应地发生了较大的变化[22,6],据此可以推断,群落集合功能特征值的变化可以代表具有不同特征值的物种的更替变化。单个物种的功能特征对生态系统的影响是通过该物种在群落中的丰富度(或生物量)而体现的,因此,在考虑物种功能特征对生态系统功能的影响时,采用群落水平功能特征指标能够更加客观地反映物种成分随环境变化而产生的作用[1],同时也使物种与群落、生态系统间建立了密切的联系[16]。
地上净初级生产力体现着草地生态系统中碳和能量的输入,是生态系统功能的重要指标[23-25]。Grime[10]的‘生物量比率理论’认为,群落中优势物种的功能特征对生态系统功能 (如生产力,分解性及养分循环)有决定性影响。本研究结果也表明,草地生态系统功能(净初级生产力)与群落中优势物种的功能特征(植株高度、相对生长率、比叶面积和叶干物质含量等)有直接或间接的关系。演替过程中由于具备不同特征的优势物种的替换结果而导致群落功能特征的变化,进而影响到净初级生产力,证明优势物种的功能特征对生态系统功能有重要影响[9],此结果可为“生物量比例理论”在植物群落演替研究中的应用提供新的依据。
本研究所选的植物特征中,植物的高度是最基本的功能特征,体现了植物空间 (地上)资源的竞争能力,群落中优势物种高度的增加,有利于光能资源的竞争[26,27],进而导致地上净初级生产力的提高(图3)。相对生长率是物种响应环境变化的关键性特征[28],该指标反应植物的生长与养分利用对策的信息,相对生长率较高的物种获取资源的能力较强,因而具有相对较高的生产力水平[16]。本研究结果也证明,优势物种的相对生长率与地上净初级生产力呈显著正相关关系 (图3)。比叶面积代表植物单位干重叶的光能截获面积,与植物的同化率密切相关[29],与植物的生存对策有密切的联系[30],也是植物重要的功能特征之一[9,31]。因而,较高的比叶面积对应着较高的生产力水平 (图3)。叶干物质含量代表叶组织密度,与植物养分保持能力密切相关[6,31]。叶干物质含量较高,植物生长较慢,具有较低的SLA值,对群落的生产力产生间接影响 (图3)。本研究中植株高度和生长率的降低伴随着比叶面积的下降和叶干物质含量的增加(图1),表明随着演替的进程,资源获取能力较强的物种逐渐被资源保存能力较强的物种所替代[6],因而导致草地生态系统的净初级生产力下降。
综上所述,高寒草甸的恢复演替对植物功能特征有重要影响,而植物的功能特征对生态系统生产力又产生直接或间接的影响。随着演替的进程,植物的株高、相对生长率和比叶面积值呈下降趋势,进而导致草地净初级生产力下降。叶干物质含量则随演替梯度的增加而增加,与地上净初级生产力相关性不明显。本研究结果表明,植物的株高、相对生长率和比叶面积对高寒草甸生态系统生产力随演替变化有一定的指示作用。在高寒草地生态系统管理和利用中,可将此特征作为预测草地系统功能响应环境变化的参考依据。
[1] Chapin F S III.Effects of plant traits on ecosystem and regional processes:a conceptural framework for prediction the consequences of global change[J].Annals of Botany,2003,91(4):455-463.
[2] Diaz S,Hodgson J G,Thompson K,etal.The plant traits that drive ecosystems:Evidence from three continents[J].Journal of Vegetation Science,2004,15(3):295-304.
[3] Hooper D U,Chapin F S III,Ewel J J,etal.Effects of biodiversity on ecosystem functioning:A consensus of current knowledge[J].Ecological Monographs,2005,75(1):3-35.
[4] Craine J M,Tilman D,Weding D,etal.Functional traits,productivity and effects on nitrogen cycling of 33 grassland species[J].Functional Ecology,2002,16(5):563-574.
[5] Díaz S,Cabido M.Vive la différence:plant functional diversity matters to ecosystem processes[J].Trends in Ecology &Evolution,2001,16(11):646-655.
[6] Garnier E,Cortez J,Billès G,etal.Plant functional markers capture ecosystem properties during secondary succession[J].E-cology,2004,85(9):2630-2637.
[7] Jiang X L,Zhang W G,Wang G.Effects of different components of diversity on productivity in artificial plant communities[J].Ecological Research,2007,22(4):629-634.
[8] Díaz S,Lavorel S,McIntyre S,etal.Plant trait responses to grazing- A global synthesis[J].Global Change Biology,2006,(13):12,1-29.
[9] Lavorel S,Garnier E.Predicting changes in community composition and ecosystem functioning from plant traits:revisiting the Holy Grail[J].Functional Ecology,2002,16:545-556.
[10] Grime J P.Benefits of plant diversity to ecosystems:immediate,filter and founder effects[J].Journal of Ecology,1998,86(6):902-906.
[11] Chapin F S III,Matson P A,Mooney H A.Principles of Terrestrial Ecosystem Ecology[M].New York,USA:Springer-Verlag,2002.
[12] Grime J P.Plant Strategies,Vegetation Processes,and Ecosystem Properties[M].UK:John Wiley and Sons Chichester,2001.
[13] Westoby M,Falster D S,Moles A T,etal.Plant ecological strategies:some leading dimensions of variation between species[J].Annual Review of Ecology,Evolution,and Systematics,2002,33(1):125-159.
[14] Garnier E,Shipley B,Roumet C,etal.A standardized protocol for the determination of specific leaf area and leaf dry matter content[J].Functional Ecology,2001,15(5):688-695.
[15] Hunt R.Plant Growth Analysis[M].London,UK:Arnold,1978.
[16] Eviner V T III,Chapin F S.Functional matrix:a conceptual framework for predicting multiple plant effects on ecosystem processes[J].Annual Review of Ecology,Evolution and Systematics,2003,34(3):455-485.
[17] Vile D,Shipley B,Garnier E.Ecosystem productivity can be predicted from potential relative growth rate and species abundance[J].Ecology Letters,2006,9(9):1061-1067.
[18] 宋晓谕,张仁懿,李新娥,等.甘南亚高山草甸弃耕演替过程中的物种多样性与生产力变化模式及相互关系研究[J].草业学报,2010,19(6):1-8.
[19] 韩立辉,尚占环,任国华,等.青藏高原“黑土滩"退化草地植物和土壤对秃斑面积变化的响应[J].草业学报,2011,20(1):1-6.
[20] Gleeson S K,Tilman D.Plant allocation,growth rate and successional status[J].Functional Ecology,1994,8(4):543-550.
[21] Reich P B,Ellsworth D S.Leaf carbon and nutrient assimilation and conservation in species of differing successional status in an oligotrophic Amazonian forest[J].Functional Ecology,1995,9(1):65-76.
[22] Garnier E,Bellmann G L A,Debain S,etal.Consistency of species ranking based on functional leaf traits[J].New Phytologist,2001,152:69-83.
[23] Mc Naughton S J,Oesterheld M,Frank D A,etal.Ecosystem-level patterns of primary productivity and herbivory in terrestrial habitats[J].Nature,1989,341:142-144.
[24] Garnier E,Lavorel S,Ansquer P,etal.Assessing the effects of land-use change on plant traits,communities and ecosystem functioning in grasslands:A standardized methodology and lessons from an application to 11 European sites[J].Annals of Botany,2007,99(5):967-985.
[25] 王长庭,龙瑞军,王根绪,等.高寒草甸群落地表植被特征与土壤理化性状、土壤微生物之间的相关性研究[J].草业学报,2010,19(6):25-34.
[26] Westoby M.A leaf-height-seed(LHS)plant ecology strategy scheme[J].Plant and Soil,1998,199(2):213-227.
[27] Westoby M,Falster D S,Moles A T,etal.Plant ecological strategies:some leading dimensions of variation between species[J].Annual Review of Ecology,Evolution,and Systematics,2002,33(1):125-159.
[28] Chapin F S III,Autumn K,Pugnaire F.Evolution of suites of traits in response to environmental stress[J].The American Naturalist,1993,142(suppl.):78-92.
[29] Reich P B,Walters M B,Ellsworth D S.Leaf life-span in relation to leaf,plant,and stand characteristics among diverse ecosystems[J].Ecological Monographs,1992,62(3):365-392.
[30] Vendramini F,Díaz S,Gurvich D E,etal.Leaf traits as indicators of resource-use strategy in floras with succulent species[J].New Phytologist,2002,154(1):147-157.
[31] Pontes L D,Soussana J F,Louault F,etal.Leaf traits affect the above-ground productivity and quality of pasture grasses[J].Functional Ecology,2007,21(5):844-853.
