粪肠球菌TLME3株AceA基因的克隆和序列分析*
2012-01-24聂鑫,高原
聂 鑫,高 原
粪肠球菌是一种新的人兽共患病病原体[1],它不但是引起医院内病人感染和死亡的主要致病菌[2],而且动物感染粪肠球菌发病和死亡的报道也越来越多[3]。猪源粪肠球菌毒力岛基因与人源相关基因、致羔羊脑炎粪肠球菌毒力因子基因片段与医学临床中某些该菌的相关基因片段同源性很高,存在着基因水平转移或某种联系[4-5]。粪肠球菌感染致死主要由其毒力因子所致。在粪肠球菌感染过程中,是由Ace介导黏附在宿主细胞外基质(ECM)蛋白上,完成定植和感染第一步的[6]。因此,Ace是粪肠球菌感染的最重要的毒力因子之一。
1 材料与方法
1.1 菌株与载体 所用粪肠球菌TLME3株为本实验室从发生败血症的雏鹅体内分离并鉴定;E.coliDH5α为本实验室保存;pMD18-T克隆载体购于大连Takara公司。
1.2 工具酶及试剂Taq酶购自Promega公司,限制性内切酶BamHⅠ和XhoⅠ、Proteinase K购于大连Takara公司,dNTPs、DNA DL2 000Marker、DNA DL10 000Marker、Solarbio凝胶回收试剂盒购自上海Solarbio生物科技公司,北京庄盟质粒小量抽提试剂盒购自北京庄盟国际生物基因科技有限公司。其它化学试剂为国产分析纯产品。
1.3 引物设计与合成 根据GenBank上已发表的粪肠球菌B-343/TX2783株(注册号为 AF260896)的 AceA基因序列,用Primer Premier 5.6.0和 Oligo 6.71软件设计1对引物,引 物 序 列 为:上 游 引 物 P1(BamH Ⅰ ):5′-CGCGGATCCAGATCACACTACT-3′,下游引物 P2(XhoⅠ):5′-,由大连 Takara公司合成。
1.4 PCR扩增 PCR反应体系:ddH2O 14.3μL,10×PCR buffer1.5μL,2.5mmol/LdNTP 2.0μL,上、下游引物各2.0 μL,模板DNA3.0μL,TaqDNA Polymeras 0.2μL,总体积25μL。PCR反应条件:95 ℃,5min;94 ℃,1min;52 ℃,1 min;72℃,1min;共30个循环;72℃延伸7min。PCR产物用10g/L琼脂糖凝胶电泳鉴定。
1.5 PCR产物的克隆和测序 用Solarbio凝胶回收试剂盒纯化回收PCR产物与pMD18-T载体连接,连接产物转化入E.coliDH5α感受态细胞,涂布于铺有IPTG(0.1mol/L)和X-Gal(0.05mol/L)的LB的平板上,37℃培养过夜。进行蓝白斑筛选,挑取生长良好的白色单个菌落进行增菌培养,用北京庄盟质粒小量抽提试剂盒提取重组质粒,进行BamH I、XhoI单、双酶切鉴定和PCR鉴定,阳性重组质粒命名为pMD18-T-AceA。筛选阳性重组质粒送大连Takara公司测序。
1.6 结构和序列分析 用Lasergene7.0及ANTHEPROT本地软件对粪肠球菌TLME3株AceA蛋白序列的组分、分子质量、滴定曲线与等电点进行预测;用Gene Runner软件,对TLME3株AceA蛋白序列一级结构中糖基化位点、磷酸化位点和硫酸化位点等修饰位点和模序进行预测;用同源建模服务器SWISS-MODEL在线软件,对TLME3株AceA蛋白的三级结构进行预测。用NCBI上BLASTn和BLASTp在线软件对粪肠球菌TLME3株AceA基因核苷酸序列及其编码氨基酸序列进行同源性搜索;用Lasergene7.0中MegAlign程序,将测定的TLME3株AceA基因核苷酸序列及其编码氨基酸序列,与用BLAST在线软件搜索得到的其他粪肠球菌AceA氨基酸序列进行多重比对,并用MEGA4.0软件Phylogeny程序 Maximum Parsimony(MP)方法生成系统进化发育树;根据序列比对结果计算TLME3株AceA氨基酸序列的变异率。
2 结 果
2.1 目的基因的扩增 以粪肠球菌TLME3株DNA为模板,用特异性引物进行PCR扩增,得到一个大小约957bp的条带,与预期结果一致,见图1。
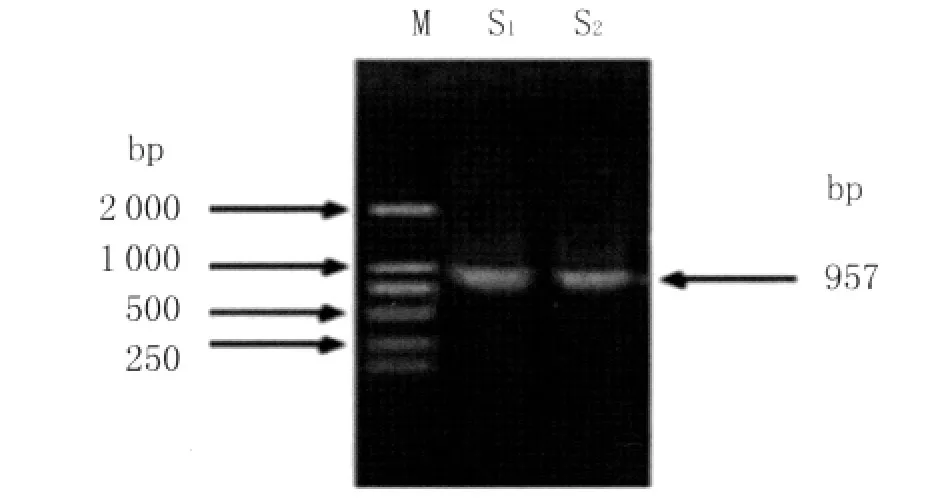
图1 粪肠球菌TLME3株AceA基因的PCR扩增M:DNA分子质量标准;S1:TLME3AceA PCR扩增产物;S2:ATCC29200AceA PCR扩增产物Fig.1 PCR amplification of AceA gene from E.faecalis strain TLME3M:DL2000DNA Marker;S1:Amplified Product of AceA from TLME3by PCR;S2:Amplified-Product of ATCC29200AceA by PCR
2.2 重组质粒的鉴定 将粪肠球菌TLME3株阳性重 组 质 粒 pMD18-T-AceA 进 行 PCR 鉴 定、BamH I单酶切鉴定和BamH I、XhoI双酶切鉴定,结果见图2。图2中3 649bp的单酶切条带是pMD18-T-AceA全长序列,2 692bp和957bp的双酶切条带分别是载体条带和目的条带,与预期结果一致,表明目的片段已经成功地与克隆载体连接。
阳性重组质粒测序结果TLME3株AceA基因核苷酸长957bp,用Lasergene7.0EditSeq程序查找的阅读框为918bp,编码306个氨基酸。将TLME3株AceA基因序列录入GenBank中,登录号为:JQ726492。
2.3 结构和序列分析
2.3.1 编码蛋白理化特性及一级和三级结构预测用Lasergene7.0对TLME3株AceA蛋白组分、分子质量、滴定曲线和等电点进行预测。
AceA含有19种氨基酸,其中Thr最多,其次为Glu和Asn,疏水性氨基酸Ala、IIe、Leu、Pro、Val和Trp共82个,占总数的26.8%;分子质量为36.09kD;滴定曲线上等电点为4.32,表明AceA是酸性蛋白。
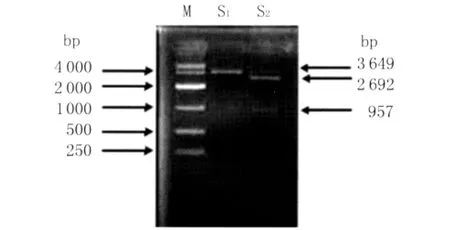
图2 重组质粒pMD18-T-AceA单、双酶切鉴定M:DNA分子质量标准;S1:重组质粒单酶切;S2:重组质粒双酶切Fig.2 Identification of pMD18-T-AceA by double restriction enzymes digestionM:DL10,000DNA Marker;S1:The product from pMD18-T-AceA digested with BamH I;S2:The product from pMD18-T-AceA digested with BamH I and XhoI
用Gene Runner软件预测的TLME3株AceA蛋白一级结构中,含有3个N糖基化位点、4个蛋白激酶C磷酸化位点、5个N肉豆蔻酰化位点和11个酪氨酸激酶Ⅱ磷酸化位点。
用同源建模服务器SWISS-MODEL在线软件预测的TLME3株AceA蛋白的三级结构,与搜索到的其他菌株AceA蛋白的3级结构同源性为98.63%,e值为5.94197e-68,可信度很大。
2.3.2 核苷酸及编码氨基酸序列同源性和变异性分析 用BLASTn搜索出58个核苷酸同源序列,其中同源性为100%的序列1个、99%的序列20个、98%的序列34个、97%的序列3个;E值全部为0。用BLASTp搜索出84个氨基酸同源序列,其中同源性为100%的序列1个、99%的序列10个、98%的序列53个、97%的序列17个,96%的序列3个;E值也全部为0。细菌分离时间为1999年-2012年,来源为医学临床和人,分离地点为美国Houston和波兰Warsaw。
用Lasergene7.0软件 MegAlign程序Jotun Hein Method方法进行核苷酸序列及其编码的氨基酸序列比对,表1是根据序列比对结果列出的TLME3株AceA氨基酸及核苷酸变异情况。

表1 粪肠球菌TLME3株AceA基因核苷酸及编码氨基酸的变异情况Tab.1 Variation situation of nucleotide and amino acid coded by AceA gene fromE.faecalis strain TLME3
用 MEGA 4.0MP方法构建的TLME3株AceA氨基酸序列系统发育进化树如图3。
3 讨 论
本试验中PCR扩增、pMD18-T-AceA重组质粒PCR鉴定、pMD18-T-AceA重组质粒单、双酶切鉴定、AceA基因序列测定等结果显示,成功克隆了粪肠球菌TLME3株AceA基因。通过核苷酸序列分析,证实插入序列与靶基因一致,阅读框正确,长度为918bp,编码306个氨基酸。为在原核细胞中表达粪肠球菌TLME3株AceA蛋白进而对其功能进行研究奠定了基础。
BLAST是由NCBI推出的搜索核苷酸和氨基酸序列数据库的最基本工具,它集速度、敏感性、弹性与统计处理的最佳结合于一身;在报告统计结果时除了显示相似性以外,也描述相似片段出现的可能性-E值;是当前最受欢迎的搜索程序之一。本研究用BLASTn搜索出的同源序列与粪肠球菌TLME3株AceA基因核苷酸序列高达97%~100%的同源性和E值全部为0的结果表明,粪肠球菌AceA基因很保守,靶序列与同源序列肯定是同源基因。58个序列中,除了3个来源不明外,其余55个均来自于医学临床。目前还没有在Gen-Bank注册的动物源粪肠球菌AceA基因核苷酸序列。
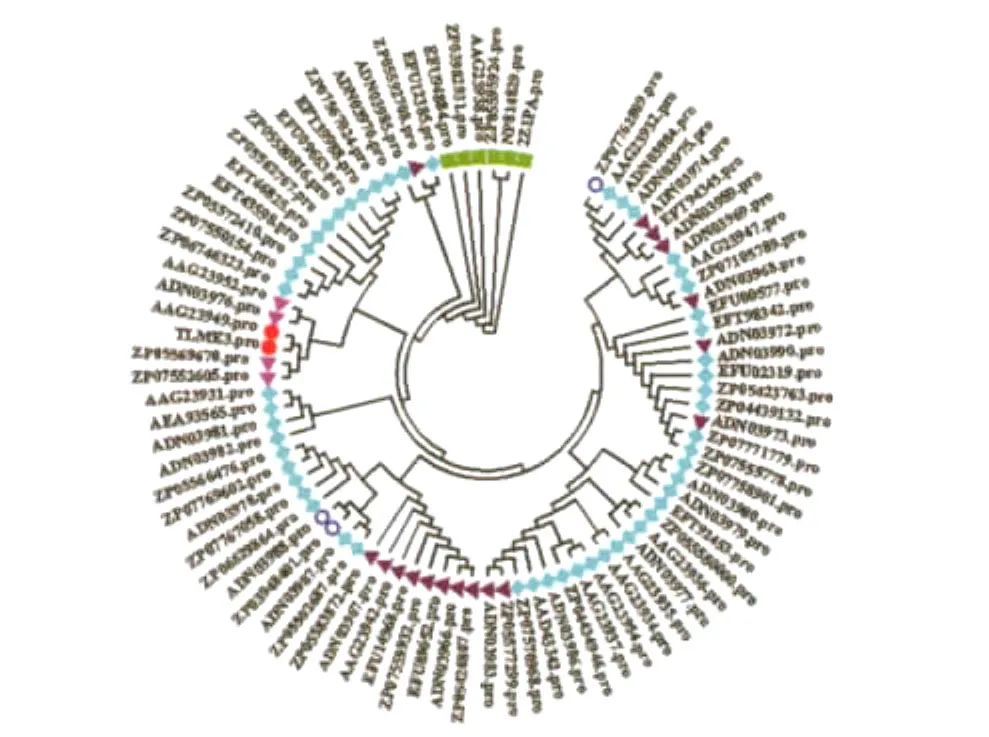
图3 TLME3株AceA氨基酸序列系统发育进化树Fig.3 Phylogenetic tree based on the protein of AceA from the strain TLME3
在系统进化发育树上,粪肠球菌TLME3株AceA与用BLASTp搜索到的84个蛋白质序列处于一个分支的2个节点上,表明它们来源于共同的祖先;与AAG23949处在同一位点上,同源性为100%;与ZP05569670、ZP07552605、ADN03976及AAG23952遗传距离最近,在一个小分支上,同源性为99%;与其余79个序列遗传距离也较近,同源性在96%~99%之间。
在本试验克隆的粪肠球菌TLME3株AceA基因阅读框核苷酸序列918个碱基中,有12个碱基变异,变异率为1.31%。其中只有6个核苷酸变异引起了编码的5个氨基酸发生了变异〔113G→T(2)和114C→T(3)是编码同一氨基酸的2个碱基〕,变异率为1.63%。在进化过程中,蛋白质序列较DNA序列更为保守,所以采用蛋白质序列进行同源性搜索更有意义。蛋白质序列搜索、系统进化发育树构建和变异率计算结果都表明,粪肠球菌AceA基因很保守,具备保守性这个动物粪肠球菌性传染病诊断及保护性抗原候选基因的关键条件。
粪肠球菌Ace是分泌在菌体外的表面蛋白,具有IgG样折叠结构,可能具有免疫活性[7]。目前国内外的试验证明了来源于人的粪肠球菌Ace蛋白对实验动物(兔、小鼠)具有免疫活性[8-10]。由于粪肠球菌TLME3株能引起雏鹅败血性疾病,造成高达30%~50%的病鹅死亡;致病性试验中,小鼠各组死亡率总和为2.33,LD50为10-1.6229[11]。如果其AceA蛋白具有免疫活性,将其作为雏鹅粪肠球菌性败血症疫苗和诊断生物制剂的候选蛋白,有重要的研究价值。
[1]Larsen J,Schoheyder HC,Lester CH,et al.Porcine-origin gentamicin resistantEnterococcusfaecalisin humans,Denmark[J].Emerg Infect Dis,2010,16(2):682-684.DOI:10.3201/eid1604.090500.
[2]Freitas AR,Tedim AP,Novais C,et al.Global spread of the hylEfm colonization virulence gene in megaplasmids of theEnterococcusfaeciumCC17polyclonal subcluster[J].Antimicrob Agents Chemother,2010,54(6):2660-2665.DOI:10.1128/AAC.00134-10.
[3]Graham JP,Price LB,Evans SL,et al.Antibiotic resistant enterococci and staphylococci isolated from flies collected near confined poultry feeding operations[J].Sci Total Environ,2009,407(8):2701-2710.DOI:10.1289/ehp.7473.
[4]Shankar N,Baghdayan AS,Willems R.Presence of pathogenieity island genes inEnterococcusfaecalisisolates from Pigs in Denmark[J].J Clin Microbiol,2006,44(11):4200-4203.DOI:10.1128/JCM.01218-06
[5]Zhou X.The finding of lamb encephalitis induced by enterococcus and studies on its pathogenic characteristics and diagnosis method[D].Ya’an:Sichuan Agriculture University,2007:89.(in Chinese)周霞.肠球菌性羔羊脑炎的发现及其病原特性和诊断方法研究[D].雅安:四川农业大学,2007:89.
[6]Singh KV,Nallapareddy SR,SillanpääJ,et al.Importance of the collagen adhesin Ace in pathogenesis and protection againstEnterococcusfaecalisexperimental endocarditis[J].PLoS Pathog,2010,6(1):e1000716.DOI:10.1371/journal.ppat.1000716
[7]Liu Q, Ponnuraj K, Xu Y. TheEnterococcusfaecalisMSCRAMMACE binds its ligand by the Collagen Hug model[J].J Biol Chem,2007,282(27):19629-19637.DOI:0.1074/jbc.M611137200
[8]Nallapareddy SR,Qin X,Weinstock GM,et al.Enterococcus faecalisadhesin,ace,mediates attachment to extracellular matrix proteins collagen type IV and laminin as well as collagen type I[J].Infect Immun,2000,68(9):5218-5224.DOI:10.1128/IAI.68.9.5218-5224.2000.
[9]Wu N.Expression and purification of ace recombinant protein ofEnterococcusfaecalisand characterization of its adherence[D].Shanghai:Nanhua University,2007:3-4.(in Chinese)武楠.粪肠球菌Ace重组蛋白的表达、纯化及黏附活性研究[D].南华大学,2007,3-4
[10]Lebreton F,Riboulet-Bisson E,Serror P,et al.ace,which Encodes an adhesin inEnterococcusfaecalis,is regulated by Ers and is involved in virulence[J].Infect Immun,2009,77(7):2832-2839.DOI:10.1128/IAI.01218-08.
[11]Di TT,Gao Y,Nie X,et al.Isolation and identification ofEnterococcusfaecaliscausing geese septicemia[J].Chin J Pre Vet Med,2012,34(3):192-196.(in Chinese)狄婷婷,高原,聂鑫,等.致鹅败血症粪肠球菌的分离鉴定[J].中国预防兽医学报,2012,34(3):192-196.DOI:10.3969/j.issn.1008-0589.2012.03.07
