社区相关耐甲氧西林金黄色葡萄球菌分子进化研究进展
2012-01-08耿文静沈叙庄
耿文静 沈叙庄
1961年,甲氧西林问世2年后,出现了第一株耐甲氧西林的金黄色葡萄球菌(methicillin-resistantStaphylococcusaureus, MRSA)。随后的20年间MRSA逐渐成为医院感染的主要病原菌,称为医院相关MRSA(hospital-associated MRSA,HA-MRSA)。20世纪90年代初期出现社区相关MRSA(community-associated MRSA,CA-MRSA)感染并逐渐增多,最近10年CA-MRSA在没有易感因素的健康人群出现,主要涉及儿童和年轻人 。
1 CA-MRSA的出现
美国CDC定义CA-MRSA为从门诊患者,或是从住院48 h内的患者中分离出来的菌株。此外,这些患者必须没有MRSA感染或定植的医疗史,在过去1年中没有住院治疗(如外科手术)、入住疗养院或透析的医疗史,并且没有永久的留置设备,如导管或其他穿通皮肤的医疗用具。无论在社区还是在医疗环境,CA-MRSA的出现都是一个严重的威胁。1993年CA-MRSA首次从澳大利亚西部偏僻社区中的土著患者中发现[1]。最近的报告表明,CA-MRSA的患病率正在增加[2]。 2003年,Salgado等[3]对住院患者和社区人群中CA-MRSA患病率的研究显示,大多数携带CA-MRSA的人至少有一项MRSA的危险因素;CA-MRSA在没有危险因素的个体中的携带率仅为0.24%。最近几年,CA-MRSA已经开始取代HA-MRSA,尤其是在CA-MRSA患病率高的地区,比如美国和中国台湾[4,5]。Moran等[6]报道,在美国11个城市的急诊科中,分离出的菌株78%是MRSA,在所有MRSA中98%与CA-MRSA常见克隆USA300有关。Gonzalez等[7]发现在美国休斯顿的一个医疗中心,60%的院内MRSA菌株与USA300克隆有关。过去的几年里,在MRSA患病率一直比较低的国家(如丹麦、挪威和荷兰)CA-MRSA的发生在一定程度上提升了MRSA的流行率,并且也发现了除USA300之外的其他克隆,CA-MRSA克隆存在着多样性[8,9](表1)。
1998年,世界范围内首次报道了儿童CA-MRSA感染[10],之后社区中健康儿童CA-MRSA感染成为世界范围内的严峻问题[11]。美国德克萨斯儿童医院在对MRSA儿童感染长达14年的回顾性研究中发现,在1990至2003年的1 002例MRSA感染的患儿中,93%(928例)是CA-MRSA。美国费城儿童医院3年的回顾性研究发现,MRSA在儿童金黄色葡萄球菌(SA)感染比例已经从15%升至40%,并且多数为皮肤与软组织感染[12]。Li等[13]研究发现,美国夏威夷CA-MRSA的流行率儿童(24%)已高于成人(21%) 。中国台湾地区报道44%儿童社区获得性SA感染为CA-MRSA。CA-MRSA菌株主要导致健康儿童青少年浅表的皮肤和软组织感染,但同时也能造成侵入性感染,导致严重的脓毒症和肺炎[14]。
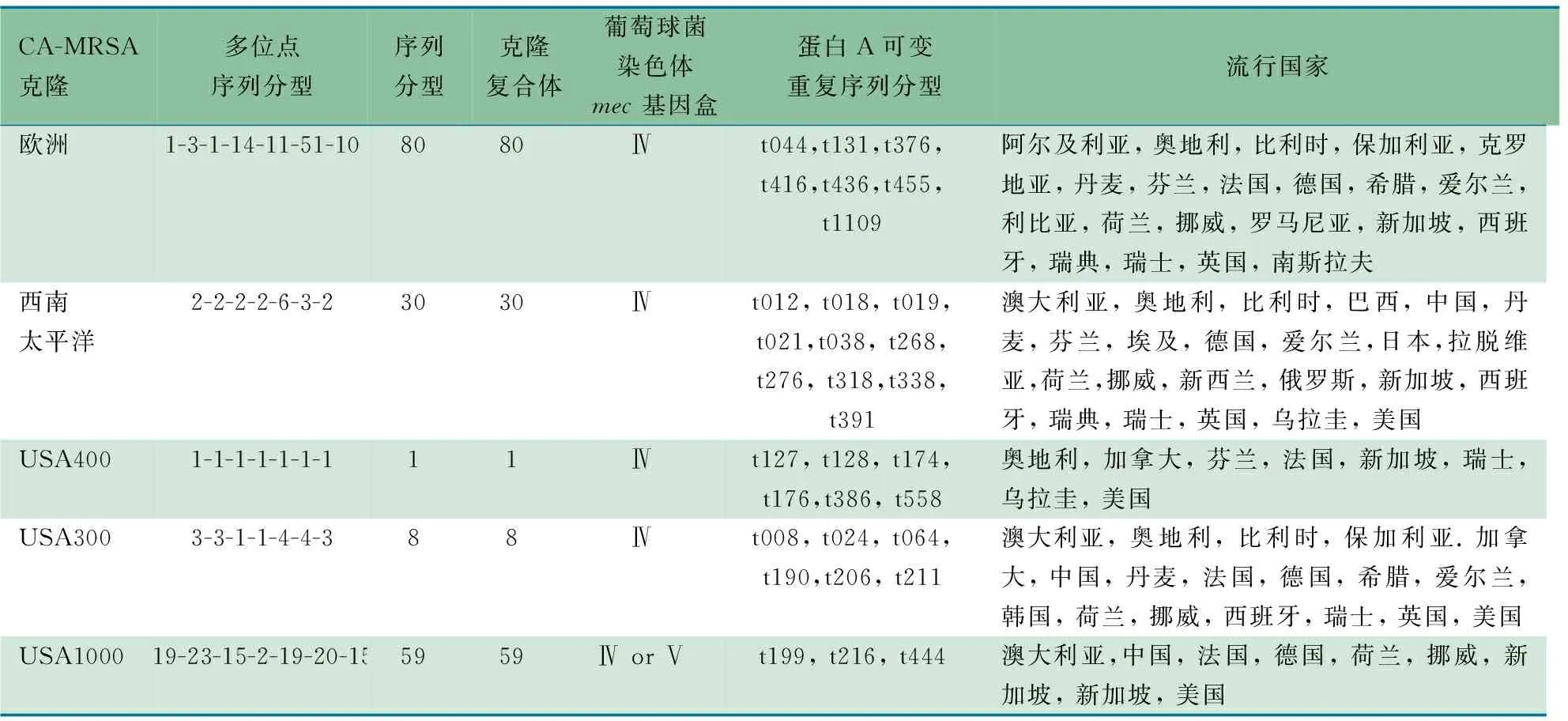
表1 世界范围内主要的CA-MRSA克隆
2 CA-MRSA的起源和进化
目前还不清楚CA-MRSA是否为社区中的甲氧西林敏感金黄色葡萄球菌(MSSA)获得SCCmec转化而来,以及CA-MRSA是否从HA-MRSA衍生而来。DeLeo等[15]认为CA-MRSA是SCCmecⅣ型被社区中MSSA获得后的新产物。日本的研究提出,20世纪60年代已经存在于日本社区中的杀白细胞毒(PVL)阳性的MSSA菌株获得SCCmecⅣ型后,便出现了PVL阳性的ST30-MRSA-Ⅳ克隆[16]。这个假设被Monecke等[17]的研究支持,SCCmec与遗传背景多样的PVL阳性的SA整合,会导致CA-MRSA的出现。然而,Aires de Sousa等[18]在CA-MRSA和 HA-MRSA菌株中观察到若干相似性,认为某些CA-MRSA菌株可能起源于医院。最近的研究显示,HA-MRSA和CA-MRSA克隆有着共同祖先。20世纪50年代,在全世界的医院和社区中出现了一种耐青霉素的SA克隆(噬菌体型80/81),但自从20世纪60年代引入耐青霉素酶的β内酰胺类抗生素后,这一克隆消失了。包含PVL的ST30 克隆再度出现,并获得SCCmecⅣ 型成为ST30-MRSA-Ⅳ 社区相关的西南太平洋克隆。ST30-MSSA也可以获得SCCmecⅡ型,可能通过几个中间的步骤,例如获得SCCmecⅣ型,成为了ST36-MRSA-Ⅱ这一流行的EMRSA-16医院相关的克隆。对从美国医疗保健设施中分离出的散发“过渡的”PVL阴性的CA-MRSA菌株进一步的研究,可能进一步阐明CA-MRSA的起源和进化[19]。
3 CA-MRSA与HA-MRSA分子特征的不同
正确区分CA-MRSA和HA-MRSA是非常重要的,两者来源不同,基因分型不同,耐药谱不同,治疗选用的抗生素也不相同。两者区分可遵循以下几条原则:①感染人群:根据美国CDC标准,CA-MRSA感染者不具有医院获得性MRSA感染者的因素,如频繁住院治疗、外科手术后、血液透析和长期护理等全身免疫功能低下的状态,而以平素身体健康的青少年为主。②耐药表型:HA-MRSA由于含有多个耐药基因,除对β内酰胺类抗生素耐药外,对氨基糖苷类、大环内酯类和喹喏酮类抗生素也不敏感。CA-MRSA除mecA基因外,一般不含有其他耐药基因,所以除对β内酰胺类抗生素耐药外,对非β内酰胺类抗生素多显示敏感。所以两者治疗选用抗生素也不相同。③基因型:采用SCCmec基因分型,结合脉冲场凝胶电泳(pulse-field gel electrophoresis, PFGE)和多位点序列分型(multilocus sequence typing,MLST)分析,显示CA-MRSA菌株的克隆型与HA-MRSA无关[20]。此外,CA-MRSA与HA-MRSA相比,有更多的克隆多样性,提示多种遗传背景的SA有可能会变成CA-MRSA[21]。CA-MRSA通常携带SCCmecⅣ、Ⅴ或Ⅶ型,而HA-MRSA的SCCmec类型主要为Ⅱ和Ⅲ型。④毒力:CA-MRSA 菌株基因序列分析中含有PVL基因,为lukS-PV和lukF-PV基因复合物,可产生PVL,相对于HA- MRSA容易丢失PVL基因而言,CA-MRSA产生的PVL致病力更强[22]。CA-MRSA 中还有葡萄球菌肠毒素基因,如seb、sec和she基因等,可产生超抗原肠毒素B和C,可使患者发生超敏反应,从而使免疫功能低下的患者出现中毒性休克综合征[23]。
4 世界范围内CA-MRSA的流行克隆
目前5种主要的PVL阳性的CA-MRSA克隆已经散布全世界。在亚洲,欧洲和美国发现ST1克隆,在欧洲和美国发现ST8克隆,在澳大利亚、欧洲和南美洲发现了ST30克隆,在亚洲和美国发现ST59克隆,在亚洲、欧洲和中东有ST80克隆。此外,已经发现有很多国际交流的国家(如游客较多的新加坡 ),CA-MRSA克隆种类最多[24]。这些CA-MRSA 克隆包含SCCmecⅣ型,ST1克隆包含Ⅳa变异体,ST80克隆包含SCCmecⅣc型。同时ST80克隆通常包含编码夫西地酸耐药的fusB基因。
除上述提到的主要CA-MRSA克隆之外,最近也发现了一些较少的PVL阳性的CA-MRSA克隆。包括散布在阿尔及利亚、法国、爱尔兰、斯洛文尼亚、西班牙和瑞士的ST5-MRSA-Ⅳ克隆,在德国、爱尔兰和荷兰发现的ST22-MRSA-Ⅳ克隆,在荷兰发现的ST37-MRSA-Ⅳ克隆,在澳大利亚广泛传播的ST93-MRSA-Ⅳ (Queensland)克隆,散布在澳大利亚、法国、希腊、瑞士和荷兰的ST377-MRSA-Ⅴ克隆,在新加坡发现的ST766-MRSA-Ⅳ克隆[25]。在有ST5遗传背景的CA-MRSA菌株中PVL的出现特别受到关注,因为ST5有大容量传播的流行性潜力。
SCCmecⅤ型存在于ST5、8、45、59、152、573和577的澳大利亚菌株、ST59的中国台湾菌株,ST8和27的芬兰菌,ST45的乌拉圭菌株、ST1、7、8、45、59、88、188、524和573的新加坡菌株,ST377的澳大利亚、荷兰、法国、希腊和瑞士菌株,ST152的科索沃菌株中。这表明包含SCCmecⅤ型的CA-MRSA菌株有不同的遗传背景。尽管包含SCCmecⅤ型的CA-MRSA菌株已经在许多国家发现,但在澳大利亚和中国台湾最流行。沈叙庄等[26]研究发现中国大陆儿童CA-MRSA分离株以CC59克隆为主,最主要的流行型是ST59-MRSA-Ⅳa-t437型,其次为ST59-MRSA-Ⅴ-t437型,与中国台湾地区类似。ST1-MRSA-Ⅳ-t127及ST1-MRSA-Ⅳ-t114型克隆也为主要的流行克隆,此克隆在美国、英国为主要流行克隆,但至今尚未在日本、中国香港和中国台湾有报道。可能提示中国大陆的儿童CA-MRSA菌株可能部分来源于距离较远的国家,如美国等[27]。
2003年荷兰观察到的MRSA克隆为ST398,大多数spa分型为t011,t034或t108,包含SCCmecⅣ或Ⅴ型,由于存在一种新的DNA甲基化酶而不能用PFGE分型[28]。在实施集中监管后,这一克隆在荷兰的患病率在2006年的年中已>21%,在2006年的年末,该克隆几乎占从荷兰医院中分离的MRSA菌株的25%,并且在奥地利和德国也观察到[29]。在丹麦的国际会议研究显示,12.5%的来自加拿大和欧洲不同国家的参与者,都携带这一MRSA克隆[30]。
5 CA-MRSA、SCCmecⅣ型和PVL的关系
SCCmecⅣ型和PVL之间是否有关系仍存在争议。有研究显示,CA-MRSA携带SCCmecⅣ型,PVL是CA-MRSA一个稳定的遗传标记[31]。CA-MRSA、SCCmecⅣ型和PVL的关系在美国Shukla 等[32]的研究中得到证实。然而,澳大利亚O'Brien 等[33]研究并未观察到在CA-MRSA、SCCmecIV型和PVL之间存在关系。而且,最近的研究显示,不足7%的CA-MRSA菌株含有PVL,提示爱尔兰PVL并不是CA-MRSA遗传标记。另外韩国的CA-MRSA主要来自于两种PVL阴性的克隆,即ST1-MRSA-Ⅳ和ST72-MRSA-Ⅳ。进一步的研究发现荷兰PVL阳性的CA-MRSA菌株包含SCCmecⅠ和Ⅲ型[34],韩国PVL阳性的CA-MRSA菌株是SCCmecⅠ和Ⅱ型[35],在阿尔及利亚发现PVL阳性的HA-MRSA菌株[35]。一般而言,40%~90%包含SCCmecⅣ型的MRSA菌株携带PVL,而<5%的包含SCCmecⅠ至Ⅲ型的MRSA菌株携带PVL[36]。
Wolter等[37]对较少见的CA-MRSA菌株进行研究,未得出PVL的等位基因变异和遗传背景之间存在联系的结论。USA300克隆另一个重要的特征是含有在其他CA-MRSA克隆中没有发现的一个PVL变异体。O'Hara 等[38]发现在CA-MRSA克隆中的PVL核苷酸序列是高度保守的,但也注意到12个核苷酸是可变的。然而,只有一个位点导致了一个氨基酸改变,如组氨酸(H变异体)变成精氨酸(R变异体)。R变异体主要发生在USA300克隆中,H变异体主要发生在其他PVL阳性的SA遗传背景中。因此推断这一氨基酸改变在PVL致病中有一定的作用。
6 CA-MRSA疾病与PVL的关系
许多研究报道PVL,SCCmecⅣ或Ⅴ型及特定的遗传背景共同组成CA-MRSA的遗传标记[37]。PVL是属于双组份毒素家族的一种SA特有的外毒素,有两种邻近和共转录的lukF-PV、lukS-PV基因编码,这两种基因各产生32和38 kDa的蛋白质[38]。像其他所有的PVL一样,PVL通过在细胞膜上产生孔来杀伤白细胞。已发现PVL阳性的SA菌与疖等原发皮肤感染和严重的坏死性肺炎有关[39]。PVL的临床重要性在1986至1998年在法国开展的SA感染的研究中进一步得到了证实,该研究报道了8例由PVL阳性的SA菌株引起的社区获得性肺炎,其中6例死亡,均为具有免疫能力的儿童和年轻人,并且每例均表现出前驱的流感样症状[40]。此外,Labandeira-Rey等[41]报道了急性肺炎鼠模型中的PVL足以引起肺组织的坏死。然而,最近也有研究显示PVL在皮肤、软组织感染和肺炎中并不起作用。Gordon等[42]用鼠脓毒症和脓肿模型来研究PVL阴性和阳性的CA-MRSA菌株的毒力,发现两类菌株有同等的毒性。同基因的PVL阴性的临床相关CA-MRSA USA300和USA400菌株在脓毒症模型中与野生型菌株均具致死性,并且也可以引起相似的皮肤感染。最近中国的一项研究发现,在有肺炎和伤口感染的老年患者中,检出5株ST398遗传背景、包含SCCmecⅢ或Ⅳ型的多重耐药、PVL阳性的CA-MRSA菌株[44]。ST398遗传背景获得PVL,向社区中个体的传播,会导致严重的医疗问题[43]。PVL能被这一克隆携带,更使得监测和限制这一克隆的传播成为一个重要的问题[44]。
总之,尽管在过去的10年里获得了许多关于MRSA克隆的认识,但仍然有许多问题没有得到回答。因此需要进一步流行病学研究,全面了解MRSA的分子进化。
[1]Udo EE, Pearman JW, Grubb WB. Genetic analysis of community isolates of methicillin-resistant Staphylococcus aureus in Western Australia.J Hosp Infect,1993, 25(2):97-108
[2]Chambers HF. The changing epidemiology of Staphylococcus aureus? Emerg Infect Dis, 2001, 7(2):178-182
[3]Salgado CD, Farr BM, Calfee DP. Community-acquired methicillin-resistant Staphylococcus aureus: a meta-analysis of prevalence and risk factors. Clin Infect Dis, 2003,36(2):131-139
[4]Otter JA, French GL. Nosocomial transmission of community-associated methicillin-resistant Staphylococcus aureus: an emerging threat. Lancet Infect Dis,2006, 6(12):753-755
[5]Seybold U, Kourbatova EV, Johnson JG, et al. Emergence of community-associated methicillin-resistant Staphylococcus aure-us USA300 genotype as a major cause of health care-associated blood stream infections. Clin Infect Dis,2006, 42(5): 647-656
[6]Moran GJ, Krishnadasan A, Gorwitz RJ, et al. Methicillin-resistant S. aureus infections among patients in the emergency department. N Engl J Med,2006, 355(7): 666-674
[7]Gonzalez BE, Mon RA.Staphylococcus aureus infections in adolescents.Adolesc Med State Art Rev, 2010,21(2):318-331
[8]Fang H, Hedin G, Li G, et al. Genetic diversity of community-associated methicillin-resistant Staphylococcus aureus in southern Stockholm, 2000-2005. Clin Microbiol Infect, 2008,14(4): 370-376
[9]Larsen AR, Bocher S, Stegger M, et al. Epidemiology of European community-associated methicillin-resistant Staphyloco-ccus aureus clonal complex 80 type IV strains isolated in Denmark from 1993 to 2004. J Clin Microbiol, 2008,46(1): 62-68
[10]Herold BC, Immergluck LC, Maranan MC, et al. Community-acquired methicillin-resistant Staphylococcus aureus in children with no identified predisposing risk. JAMA, 1998,279(8):593-598
[11]David MZ, Daum RS. Community-associated methicillin-resistant Staphylococcus aureus: epidemiology and clinical consequences of an emerging epidemic. Clin Microbiol Rev, 2010, 23(10):616-687
[12]Filice G, Nyman J, Lexau C, et al. Excess costs and utilization associated with methicillin resistance for patients with Staphylo-coccus aureus infection. Infect Control Hosp Epidemiol, 2010,31(4): 365-373
[13]Li S, Skov RL, Han X, et al. Novel types of staphylococcal cassette chromosome mec elements identified in clonal complex 398 methicillin-resistant Staphylococcus aureus strains. Antimicrob Agents Chemother, 2011,55(6):3046-3050
[14]Gould IM, Cauda R, Esposito S, et al. Management of serious meticillin-resistant Staphylo coccus aureus infections: what are the limits? Int J Antimicrob Agents, 2011,37(3):202-209
[15] DeLeo F, Otto M, Kreiswirth B, et al. Community associated meticillin-resistant Staphyl ococcus aureus. Lancet, 2010, 375(9725): 1557-1568
[16] McConeghy K, Mikolich D, LaPlante K. Agents for the decolonization of methicillin-resistant Staphylococcus aureus. Pharmacotherapy, 2009, 29(3): 263-280
[17]Monecke S, Engelmann I, Archambault M, et al.Distribution of SCCmec-associated pheno-soluble modulin in staphylococci.Mol Cell Probes, 2012,26(2):99-103
[18]Aires-de-Sousa M, Correia B, de Lencastre H.Changing patterns in frequency of recovery of five methicillin-resistant Staphylococcus aureus clones in Portuguese hospitals: survei-llance over a 16-year period.J Clin Microbiol,2008,46(9):2912-2917
[19]David MZ, Daum RS. Community-associated methicillin-resistant Staphylococcus aureus: epidemiology and clinical consequences of an emerging epidemic. Clin Microbiol Rev, 2010, 23(3):616-687
[20]Garau J, Bouza E, Chastre J, et al. Management of methicillin-resistant Staphylococcus aureus infections. Clin Microbiol Infect, 2009,15(2):125-136
[21] Loffler B, Hussain M, Grundmeier M, et al. Staphylococcus aureus Panton-Valentine leukocidin is a very potent cytotoxic factor for human neutrophils. PLoS Pathog, 2010, 6:e1000715
[22]Lipinska U, Hermans K, Meulemans L,et al. Panton-Valentine leukocidin does play a role in the early stage of Staphylococcus aureus skin infections: a rabbit model. PLoS One, 2011, 6: e22864
[23]Huang YC, Ho CF, Chen CJ, et al. Comparative molecular analysis of community-associated and healthcare-associated methicillin-resistant Staphylococcus aureus isolates from children in northern Taiwan. Clin Microbiol Infect, 2008, 14(12): 1167-1172
[24] Ruud H, Deurenberg,Ellen E, et al. The molecular evolution of hospital- and community-associated methicillin-resistant Staphylococcus aureus. Current Molecular Medicine 2009, 9:100-115
[25]Milheirico C, Oliveira DC, de Lencastre H. Update to the multiplex PCR strategy for assignment of mec element types in Staphylococcus aureus. Antimicrob Agents Chemother, 2007, 51(9): 3374-3377
[26]Geng W, Yang Y,Gen D, et al. Molecular characteristics of community-acquired, methicillin -resistant Staphylococcus aureus isolated from Chinese children. FEMS Immunol Med Microbiol, 2010,58(3):356-362
[27]Jansen WT, Beitsma MM, Koeman CJ, et al. Novel mobile variants of staphylococcal cassette chromosome mec in Staphy-lococcus aureus. Antimicrob Agents Chemother, 2006, 50(6):2072-2078
[28]Hallin M, Deplano A, Denis O, et al . Validation of pulsed-field gel electrophoresis and spa typing for long-term, nationwide epidemiological surveillance studies of Staphylococcus aureus infections. J Clin Microbiol,2007, 45(1): 127-133
[29]van Loo I, Huijsdens X, Tiemersma E,et al. Emergence of methicillin-resistant Staphylococcus aureus of animal origin in humans. Emerg Infect Dis, 2007, 13(12): 1834-1839
[30]Witte W, Strommenger B, Stanek C, et al . Methicillin-resistant Staphylococcus aureus ST398 in humans and animals, Central Europe. Emerg Infect Dis,2007, 13(2): 255-258
[31] Wulf MW, Sorum M, van Nes A, et al. Prevalence of methicillin-resistant Staphylococcus aureus among veterinarians: an international study. Clin Microbiol Infect, 2008, 14(1):29-34
[32]Shukla SK, Stemper ME, Ramaswamy SV, et al. Molecular characteristics of nosocomial and Native American community-associated methicillin-resistant Staphylococcus aureus clones from rural Wisconsin.J Clin Microbiol,2004, 42(8): 3752-3757
[33]O'Brien FG, Lim TT, Chong FN, et al. Diversity among community isolates of methicillin-resistant Staphylococcus aureus in Australia. J Clin Microbiol,2004, 42(7):3185-3190
[34]Wannet WJ, Spalburg E, Heck ME, et al.Emergence of virulent methicillin-resistant Staphylococcus aureus strains carrying Panton-Valentine leucocidin genes in The Netherlands.J Clin Microbiol.,2005,43(7):3341-3345
[35]Park C, Lee DG, Kim SW, et al. Predominance of community-associated methicillin- resistant Staphylococcus aureus strains carrying staphylococcal chromosome cassette mec type IVA in South Korea. J Clin Microbiol,2007,45(12): 4021-4026
[36] Jeong HY, Lee JE, Choi BK, et al . Molecular epidemiology of community-associated antimicrobial-resistant Staphylococcus aureus in Seoul, Korea (2003): pervasiveness of multidrug-resistant SCCmec type II methicillin-resistant S. aureus. Microb Drug Resist,2007, 13(3): 178-185
[37]Wolter DJ, Tenover FC, Goering RV, et al. A geographic variant of the Staphylococcus aureus Panton-Valentine leuko-cidin toxin and the origin of community-associated methicillin -resistant S. aureus USA300. Clin Microbiol Infect,2007, 13: 827-830
[38]O'Hara FP, Guex N, Word JM, et al. A geographic variant of the Staphylococcus aureus Panton-Valentine leukocidin toxin and the origin of community-associated methicillin-resistant S. aureus USA300. Infect Dis, 2008, 197(2):187-194
[39]Tristan A, Bes M, Meugnier H, et al. Global distribution of Panton-Valentine leukocidin-positive methicillin-resistant Staph-ylococcus aureus, 2006. Emerg Infect Dis, 2007, 13(4): 594-600
[40] Gillet Y, Issartel B, Vanhems P, et al. Association between Staphylococcus aureus strains carrying gene for Panton-Valentine leukocidin and highly lethal necrotising pneumonia in young immunocompetent patients. Lancet,2002, 359(9308): 753-759
[41] Labandeira-Rey M, Couzon F, Boisset S, et al . Staphylococcus aureus Panton-Valentine leukocidin causes necrotizing pneu-monia. Science, 2007, 315(5815): 1130-1133
[42]Gordon RJ, Lowy FD.Pathogenesis of methicillin-resistant Staphylococcus aureus infection.Clin Infect Dis,2008,46(S5):350-359
[43]Li M, Cheung GY, et al, Comparative analysis of virulence and toxin expression of global community-associated methicillin-resistant Staphylococcus aureus strains. J Infect Dis,2010,202(12):1866-1876
[44] Walkey AJ, O'Donnell MR, Wiener RS. Linezolid vs glycopeptide antibiotics for the treatment of suspected methicillin-resistant Staphylococcus aureus nosocomial pneu-monia: a meta-analysis of randomized controlled trials. Chest, 2011, 139(5):1148-1155
