豌豆属种质资源遗传多样性的ISSR分析
2012-01-02曾亮李敏权杨晓明
曾亮,李敏权,*,杨晓明
(1.甘肃农业大学草业学院,甘肃 兰州730070;2.甘肃省农业科学院,甘肃 兰州730070)
*豌豆属(Pisum)是豆科蚕豆族的一年或越年生草本植物,是粮菜饲兼用作物,喜冷冻湿润气候,幼苗能耐5℃低温,生长期适温12~16℃,结荚期适温15~20℃,具有耐寒、耐旱、耐瘠等特点,因其适应性很强,在全世界的地理分布很广[1]。中国是世界第二大豌豆主产国,广阔的地理分布范围和多样化的生境造就了丰富的豌豆种质资源,形成了豌豆的遗传多样性[2]。分析豌豆的遗传结构和遗传多样性,不仅有利于豌豆种质资源的合理保存、优异豌豆种质资源的挖掘与创新,而且对豌豆生产和育种也具有重要的指导意义。
ISSR(inter-simple sequence repeats)是一种建立在PCR反应基础上的DNA分子标记技术,由Zietkiewicz等[3]于1994年创建。该技术克服了 RFLP(restriction fragment length polymorphism)、SSR(simple sequence repeat)、RAPD(random amplified polymorphic DNA)、AFLP(amplification fragment length polymorphism)等标记技术的一些局限性,具有无需预知基因组背景信息、DNA样品用量少、信息量大、操作简单、重复性好等优点,现已被广泛应用于植物的品种鉴定、基因定位、遗传作图、进化和系统发育等方面的研究[4,5]。
对于豌豆属种质资源遗传多样性的研究,以往主要是通过形态性状和细胞学特征、SSR、AFLP、RAPD技术等进行分析,如宗绪晓等[6]利用SSR标记分析了国内豌豆地方品种的遗传多样性,21对SSR引物共扩增出104个等位变异,有效等位变异为62.52%;Hoey等[7]利用同工酶和RAPD标记方法研究发现野生型和栽培型豌豆品种间有明显差别;Samec和Nasinec[8]及Simioniuc等[9]利用RAPD和AFLP标记发现草料豌豆和粒用饲料豌豆有明显差异。但是侧重于豌豆属植物ISSR分子标记的研究尚罕见报道。本研究应用ISSR标记技术,从DNA水平上探讨了来自国内外的73份豌豆品种种质资源的遗传多样性与亲缘关系,为有效利用DNA标记评价豌豆种质资源亲缘关系,保护和合理利用种质资源以及加快新品种选育提供科学依据。
1 材料与方法
1.1 供试材料
73份豌豆材料为甘肃省农业科学院作物研究所多年来从全国各地引进和选育品种(表1),于2011年3月种植于甘肃省农业科学院秦王川试验基地。该试验地位于甘肃省兰州市永登县秦川镇,属温带大陆性季风气候,平均海拔2 100m,年降水量250~300mm。于5月采摘豌豆新鲜幼嫩叶,放入冰盒,带回实验室置于-80℃超低温冰箱保存以备DNA提取。

表1 供试材料Table 1 Experiment materials
1.2 DNA提取和定量
每份材料取10~15个植株上的新鲜叶片,DNA提取采用改进CTAB法[10];用0.8%的琼脂糖电泳检测DNA质量;将所得的DNA提取液于紫外可见分光光度计上测定波长260和280nm处的吸光度OD260和OD280,计算OD260/OD280的值以检测DNA样品纯度,检测比值为1.8~1.9;用0.1倍TE将DNA稀释至10ng/μL,置于-20℃冰箱保存备用。
1.3 ISSR引物
选用加拿大哥伦比亚大学(University of British Colunbia Biotechnology,UBC)公布的100对ISSR引物(UBC801-UBC900),由上海生工生物技术公司合成。
1.4 PCR扩增及检测
利用优化的PCR反应体系从100条ISSR引物中筛选出11条对全部DNA样品均有清晰、稳定扩增产物且条带清晰、重复性好的引物进行ISSR-PCR扩增,每条引物3次重复。20μL反应体系中各反应物含量为:15 ng模板DNA,0.2mmol/L dNTP,0.5μmol/L ISSR引物,1UTaq DNA聚合酶,2μL 10×PCR Buffer。PCR扩增程序为:94℃充分变性3min,然后进行以下34个循环:94℃变性30s,50℃退火30s,72℃延伸45s,最后于72℃延伸10min,4℃终止反应保存。
扩增产物在2%(W/V)的琼脂糖凝胶(含EB 0.5μg/mL)、1×TAE缓冲液中电泳分离(5V/cm),用2 000 bp DNA Ladder作分子量标准,在紫外凝胶成像系统下拍照记录。
1.5 数据处理与分析
同一引物扩增的电泳迁移率一致的条带被认为具有同源性,选择清晰的谱带进行统计,扩增产物在相同迁移位置有带赋值为1,无带赋值为0,建立原始数据矩阵。利用Popgen 32软件计算ISSR扩增引物的Shannon多样性信息指数,多态位点数,多态位点比率和有效等位基因数。采用NTSYS-PC 2.1软件的SAHN程序和GS值按非加权配对算术平均法(UPGMA)计算各豌豆品种间的相似系数并进行聚类分析构建聚类图。
2 结果与分析
2.1 引物筛选
从100条ISSR引物中筛选出11条多态性丰富、条带清晰且重复性较好的引物,用于供试73个豌豆材料的PCR扩增。并根据各引物的特性和具体的扩增反应对照,确定了各引物的最佳退火温度(表2)。
2.2 ISSR扩增结果及多态性分析
11条引物对73份豌豆材料共扩增出91条带,大小为200~2 000bp(图1),其中多态性条带为78条,平均多态性比率达到86.4%。每个引物可扩增出2~12条多态性条带,平均产生7条多态性谱带。其中引物 UBC818、825、836、840的多态性信息含量达到了100%。这11条引物全部是二核苷酸重复序列类型的引物,其中(GA)n有3条,(CA)n有1条,(AC)n有4条,(AG)n有2条,(CT)n有1条,说明豌豆属基因组中存在大量的(GA)、(AC)、(AG)等二核苷酸重复序列。Shannon多样性信息指数评价样品遗传多样性的结果表明,平均多样性指数(I)为0.420 2,每个位点的有效等位基因数(Ne)为1.451 8(表3),表明豌豆材料间具有较高的多态性,遗传多样性较丰富。

表2 ISSR分析的引物序列Table 2 Primer sequences used in ISSR analysis
2.3 豌豆品种间遗传相似性与差异性分析
根据扩增产物的电泳结果,通过NTSYS-PC 2.1软件对73份豌豆材料的遗传相似系数(genetic similarity)和遗传距离(genetic distance)进行计算,结果表明,73份豌豆材料间的遗传相似系数为0.406 5~0.934 0,遗传距离变幅范围为0.059 2~0.987 4cm。其中,L1417和德豌之间的遗传相似系数最小(0.406 5),遗传距离最大(0.987 4cm),说明供试材料间二者亲缘关系最远;武威珍珠绿和珍珠绿之间的遗传相似系数最大(0.934 0),遗传距离最小(0.059 2cm),说明供试材料间二者亲缘关系最近。在73份豌豆材料中来自于相同或相近地理位置的材料间遗传相似系数值较大,亲缘关系较近,而地理位置或生境相差较大的材料遗传距离值较大,亲缘关系较远。整体看来,豌豆种质资源间的遗传基础相对较宽,存在较大的遗传变异性。

表3 73份豌豆材料ISSR标记扩增结果Table 3 The result of ISSR amplification of 73pea

图1 引物UBC842对部分豌豆材料基因组DNA的PCR扩增结果Fig.1 Result of PCR amplication using primer UBC842of pea
2.4 豌豆品种间的ISSR聚类分析
利用UPGMA法对73份豌豆材料进行UPGMA聚类分析,绘出树状聚类图(图2)。在GS≈0.52处,可以把73份豌豆品种划分为5类:第1类共8份材料,其中包括来自新西兰的3份材料和国内的5份材料;第2类共13份材料,主要是来自加拿大等地的6份国外材料和7份国内材料;第3类包含的种质资源最多,共37份材料,其中绝大部分是来自我国西北和华北的豌豆种质资源;17号(X9002)与其余材料差异明显,单独聚为1类(第4类);第5类的14份材料包括13份来自我国长江中下游及西南地区和1份来自荷兰的豌豆种质资源。从聚类结果可以看出,材料地理来源对系统聚类结果影响较大,各来源地材料在聚类图中既有区别又有交叉,说明材料间形态性状变异有一定的连续性。具有相同生长习性且地理来源相近的豌豆种质资源聚为一类,聚类结果与豌豆地理来源、生长习性和生态分布密切关联。
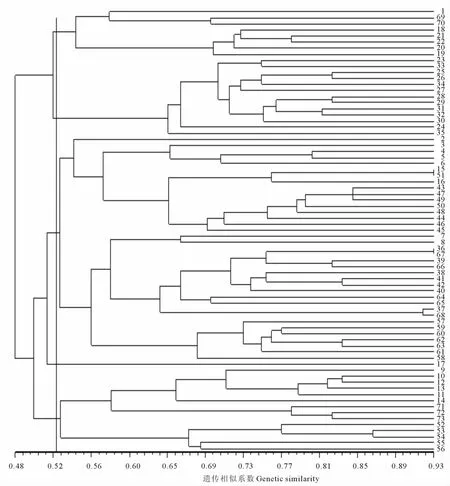
图2 基于ISSR标记的73份豌豆材料聚类图Fig.2 Dendrogram of cluster analysis based on ISSR of 73pea
3 讨论
本研究采用ISSR标记对来自国内外的豌豆属植物资源进行了综合分析,获得了较为满意的结果。由于ISSR标记具有无需预知基因组背景信息、信息量丰富、操作简单、重复性好等诸多优点,因而已被广泛应用于植物遗传多样性的研究中。胡甦等[11]采用正交设计直观分析法对影响三角紫叶酢浆草(Oxalistriangularis)ISSRPCR反应较大的5个因素在5个水平上进行了优化,并由此建立了适合三角紫叶酢浆草的ISSR-PCR反应体系。冯亮亮等[12]在对甘肃地区红砂(Reaumuriasoongorica)遗传多样性的ISSR分析表明,12条引物检测到69个位点,其中多态性位点60个,多态位点比率达到86.96%,说明ISSR标记具有较高的多态性检测水平。李鸿雁等[13]应用ISSR分子标记对分布于内蒙古不同地区的扁蓿豆(Medicagoruthenica)进行了遗传多样性研究,结果表明,15个引物共扩增出363个位点,聚类结果显示,生态地理条件相似的种群优先聚类。王照兰等[14]对扁蓿豆4个不同品系的ISSR遗传多样性分析表明,7个有效引物共扩增出73个位点,平均每条引物检测出10.4个位点,多态位点百分率达到94.5%。耿立格等[15]对河北省的绿子叶黑豆(Glycinemax)种质资源的ISSR遗传多样性分析表明,7个ISSR引物共检测出60个等位变异,平均每个位点8.6个变异,变异幅度为5~17个,聚类结果显示,类群与品种来源地无关。卢萍等[16]应用ISSR标记对小花棘豆(Oxytropisglabra)的6个地理种群进行的遗传多样性分析表明,14个引物共扩增出327个位点,多态位点比率在70%左右。从前人的研究结果可以看出,ISSR分子标记技术是有效揭示品种遗传多样性的一种手段。
宗绪晓等[17]对103份豌豆品种的SSR标记遗传多样性分析的结果表明,平均每对引物可以扩增出4.95个等位变异,多态性比率为65.56%,UPGMA聚类将供试豌豆聚成3类,生境和表型相近的种群基本聚为一类。陈永霞等[18]利用EST-SSR标记对西南扁穗牛鞭草(Hemarthriacompressa)的遗传多样性分析表明,23对引物共扩增出323条带,多态性比率80.4%,聚类分析显示,各供试材料间的聚类与其地理来源及形态特征具有一定的相关性。王海飞等[19]利用ISSR标记对527份蚕豆资源的遗传多样性分析表明,平均每对引物扩增条带数为25,多态性比率为96%,聚类结果表明,生态地理条件相似的种群优先聚集。本研究通过ISSR标记对豌豆进行遗传多样性分析表明,11条ISSR引物共扩增出91条条带,多态性比率为86.4%,这与后者的研究结果较相近,其引物扩增的条带数和多态性比率都高于用SSR分析的结果,说明ISSR标记对豌豆种质资源遗传多样性的分析更精确和可靠。聚类分析结果与前人研究结果相似,基本符合地理来源相近的居群聚为一类。
材料间遗传差异、遗传关系和遗传多样性评价对育种具有十分重要的意义[20,21]。应用分子标记技术研究遗传关系时,引物数目和标记位点数会对结果产生影响,一般来说,所用引物数目越多、检测的位点数越多,所提供的信息就越可靠。但在实际应用中,由于受到生物自身情况和时间、经费等条件的制约,可供分析利用的引物数目和位点数都是有限的。有研究表明,所分析的位点数与所能提供信息的可靠性之间具有明显的关系,当位点数超过70个时,所提供的信息可靠性趋于稳定[22]。本试验所筛选出的11个ISSR引物对73份豌豆材料共检测到91个位点,其中多态性位点78个,因此能提供可靠的信息。
从73份豌豆材料的聚类图可以看出在GS约为0.52处将73份材料聚为5类,聚类基本符合地理来源相近的居群聚为一类。但也有地理来源不同的品种被归入一类,产生这种结果的原因可能有2个方面:1)地理来源相同的材料虽然来自于同一环境,但是由于选择方向的不同和选用育种材料的错综复杂,也可能形成遗传差异较大的类型,因此出现地理来源相同的品种被归入不同类群;2)受环境饰变作用的影响,物种在长期适应环境的过程中会出现某些性状趋同的现象,造成不同物种性状间的交叉。
4 结论
通过ISSR标记技术对73份豌豆材料的遗传多样性分析表明,100个ISSR引物中共筛选出11个多态性明显、条带清晰、反应稳定的引物,多态性比率为86.4%。Shannon多样性指数平均为0.420 2,品种间遗传相似系数变幅范围为0.406 5~0.934 0,表现出丰富的遗传多样性。利用UPGMA聚类分析,将73份材料划分为5类,聚类结果呈一定的地域性分布规律。
[1]贺晨帮,宗绪晓.豌豆种质资源形态标记遗传多样性分析[J].植物遗传资源学报,2011,12(1):42-48.
[2]宗绪晓,关建平,王述民,等.国外栽培豌豆遗传多样性分析及核心种质构建[J].作物学报,2008,34(9):1518-1528.
[3]Zietkiewicz E,Rafalski A,Labuda D.Genome fingerprinting by simple sequence repeat(SSR)-anchored polymerase chain reaction amplification[J].Genomics,1994,20:176-183.
[4]王瑜,袁庆华.紫花苜蓿ISSR-PCR反应体系的建立与优化[J].草地学报,2007,15(3):212-215.
[5]Godwin I D,Aitken E A,Smith L W.Application of intersimple sequence repeat(ISSR)markers to plant genetics[J].Electrophoresis,1997,18:1524-1528.
[6]宗绪晓,关建平,王述民,等.中国豌豆地方品种SSR标记遗传多样性分析[J].作物学报,2008,34(8):1330-1338.
[7]Hoey B K,Growe K R,Jones V M,etal.A phylogenetic analysis ofPisumbased on morphological characters,allozyme and RAPD markers[J].Theoretical and Applied Genetics,1996,92:92-100.
[8]Samec P,Nasinec V.Detection of DNA polymorphism among pea cultivars using RAPD technique[J].Biologia Plantarum,1995,37:321-327.
[9]Simioniuc D,Uptmoor R,Friedt W,etal.Diversity and relationships among pea cultivars revealed by RAPDs and AFLPs[J].Plant Breed,2002,121:429-435.
[10]顾红雅,瞿礼嘉.植物分子生物学实验手册[M].北京:高等教育出版社,1998.
[11]胡甦,王永清,陶炼,等.三角紫叶酢浆草ISSR反应体系的建立与优化[J].草业学报,2011,20(5):142-150.
[12]冯亮亮,唐红,李毅,等.甘肃红砂不同种群遗传多样性的ISSR分析[J].草业学报,2011,20(1):125-130.
[13]李鸿雁,李志勇,王小丽,等.内蒙古扁蓿豆遗传多样性的ISSR分析[J].西北植物学报,2011,31(1):52-56.
[14]王照兰,杨持,赵丽丽,等.扁蓿豆不同品系ISSR标记遗传差异和遗传多样性[J].中国草地学报,2010,32(1):11-17.
[15]耿立格,王丽娜,张磊,等.河北省绿子叶黑豆种质资源表现型和ISSR标记遗传多样性分析[J].植物遗传资源学报,2010,11(3):266-270.
[16]卢萍,赵萌莉,韩国栋,等.内蒙古小花棘豆遗传多样性的ISSR分析[J].西北植物学报,2007,27(6):1102-1107.
[17]宗绪晓,Ford R,Redden R,等.豌豆属SSR标记遗传多样性结构鉴别与分析[J].中国农业科学,2009,42(1):36-46.
[18]陈永霞,张新全,谢文刚,等.利用EST-SSR标记分析西南扁穗牛鞭草种质的遗传多样性[J].草业学报,2011,20(6):245-253.
[19]王海飞,关建平,马钰,等.中国蚕豆种质资源ISSR标记遗传多样性分析[J].作物学报,2011,37(4):595-602.
[20]刘忠辉,刘国道,黄必志,等.云南猪屎豆属遗传多样性的ISSR分析[J].草业学报,2009,18(5):184-191.
[21]范彦,李芳,张新全,等.扁穗牛鞭草种质遗传多样性的ISSR分析[J].草业学报,2007,16(4):76-81.
[22]周泽扬,夏庆友,鲁成,等.分子系统生物学研究中分子位点数与遗传差异信息可靠性的关系[J].遗传,1998,20(5):12-15.
