青藏高原东北边缘地区高寒草甸群落特征对封育的响应
2012-01-02苗福泓郭雅婧缪鹏飞郭正刚沈禹颖
苗福泓,郭雅婧,缪鹏飞,郭正刚,沈禹颖
(草地农业生态系统国家重点实验室 兰州大学草地农业科技学院,甘肃 兰州730000)
*高寒草地是青藏高原植被的主体,其不仅孕育着众多的土著生物和抗逆种质资源[1],而且是藏族牧民从事草地畜牧业生产的物质基础,更是江河源区或国家陆地边界分布区,因此,高寒草地生态系统的健康不仅事关当地食物安全,而且事关我国生态安全、民族团结和社会稳定。但长期的不合理利用导致草地群落组分变化[2],群落组分变化改变群落结构,群落结构变化迫使高寒草地生态系统功能发生调整[3],从而影响整个区域生态过程,对本区和毗邻地区生物以及生物与环境相互作用组成的生态系统产生深刻影响[4]。为应对高寒草地普遍退化所引发的一系列生态问题,国家相继启动了退牧还草和牧区移民定居工程,旨在恢复退化高寒草地的生态、生产和生计功能,其中,封育是上述生态工程常用的,也是主要的措施之一,被广泛大面积的应用[5,6]。已有研究表明,封育能够明显促进退化草地的恢复和休憩,增加了美国加利福尼亚州和我国内蒙古及天山草地的盖度、高度、地上生物量[7-9]。但近期研究表明,长期封育不仅隔离了部分野生动物迁徙的通道,影响野生动物繁殖[10],而且加重了当前未围栏区或放牧区的放牧压力,使未围栏区或放牧区草地在高强度践踏和采食干扰下加速退化[11],更为重要的是忽略了高寒草地的资源属性[12],草地与其他植被生态系统的区别之一就是放牧,放牧家畜通过利用饲用植物而发生经济效益。长期封育本质上违背了封育用于退化草地恢复和休憩的初衷,因此,量化封育时期成为评估退牧还草生态工程效益和实施期限的重要科学问题。
高寒草甸是高寒草地的主要组分之一,主要分布于我国东北、新疆天山和青藏高原,其植物群落组成主要为冷中生的多年生草本植物[13],以莎草科和禾本科为主,生长密集,植株低矮,具有强大的水源涵养功能[14],成为我国围栏封育恢复生态的重点区域。由于缺乏长期监测封育对高寒草甸植物群落的影响,目前究竟封育多长时间高寒草甸就能休养生息,可以合理利用尚无定论。
本研究通过监测青藏高原东北缘天祝地区高寒草甸群落组分、高度、盖度和生物量对封育年限的响应,诠释围栏年限对高寒草甸群落特征不同指标影响的规律性和趋同性,确定适宜的封育时间,为合理退牧和资源可持续利用提供科学依据。
1 材料与方法
1.1 研究区自然概况
甘肃省天祝藏族自治县位于青藏高原东北边缘地带,年均温-0.2~1.3℃,最高气温26℃,最低气温-30℃,年降水量266~630mm,≥0℃年积温1 327.7~1 738.2℃,海拔1 960~4 878m,属高原气候类型,土壤主要为山地栗钙土和山地黑钙土。高寒草甸是天祝县的主要草地类型,是当地退牧还草的重点草地类型。目前退化严重,牧草生长高度降低,植被覆盖度减小,优良牧草比例减少,杂类草趁势增多,毒害草大量蔓延,可食牧草产量逐渐下降[15]。
1.2 样地设置与调查
样地设置在抓喜秀龙乡南泥沟八支沟(102°47′13″E,37°10′37″N)的暖季牧场,2003年4月放牧前建立50 m×50m围栏样地3块,并在距每个围栏样地100m的地方标记50m×50m对照样地,对照样地仅在4个角用小木桩标记,不作任何处理,为当地自然放牧管理状态。然后每年8月中旬做样方调查,截至2008年。
每个样地内随机设置1m×1m的样方4个,保证样方离样地边缘的距离大于2m,记录每个样方内植物种类、高度和盖度。群落内每个物种的高度测定采用随机测定30株的方法,如果不足30株,则全部测定,群落内物种盖度测定采用针刺法,测定群落草丛高度和群落盖度,将样方内植物分种离地表1.3cm处收割,85℃烘干至恒重,称干重。
1.3 指标计算
选用Shannon-Wiener指数(H)、Pielou指数(J)和丰富度指数(S)测度物种多样性[16]。
物种重要值(Ni)=(相对盖度+相对干重+相对高度)/3,相对重要值(Pi)=Ni/∑Ni。
丰富度指数采用样方内出现的物种数(S)表示。

式中,Pi为种i的相对重要值,Ni为种i的绝对重要值,S为种i所在样方中的物种数目。
1.4 数据分析
用Excel进行数据的初步整理,SPSS软件包中的单因子方差分析(ANOVA)和最小显著差数法(LSD)对各指标间进行比较和差异显著性检验。
2 结果与分析
2.1 封育对物种组成的影响
连续6年封育对高寒草甸优势种没有显著的影响,表现为6年内封育和未封育群落内高寒草甸群落优势种均为线叶嵩草(Kobresiacapillifolia),但伴生种发生了明显变化,未封育草甸群落主要伴生种从2003年的球花蓼(Polygonumsphaerostachyum),高山唐松草(Thalictrumalpinum),珠芽蓼(P.viviparum)演变为2008年的苔草(Carextristachya),披碱草(Elymusnutans),洽草(Koeleriacristata),而球花蓼和珠芽蓼在高寒草甸群落中地位相对减弱,高山唐松草退出群落,苔草、披碱草、洽草在高寒草甸群落中地位逐渐增强。封育草甸群落主要伴生种从2003年的珠芽蓼,藏嵩草(K.tibetica),球花蒿(Artemisiasmithii),演变为2008年的珠芽蓼,藏异燕麦(Helictotrichontibeticum),矮生嵩草(K.humilis),其中封育使球花蒿和藏嵩草在高寒草甸群落中地位减弱,而矮生嵩草和藏异燕麦在高寒草甸群落中地位逐渐增强。
从2003-2008年,封育条件下,高寒草甸群落内物种数表现为初始显著下降,然后显著快速上升,最后显著下降的变化趋势(P<0.05)(表1),物种数最多年份出现在封育4年的2006年,而未封育条件下的高寒草甸群落物种数随着监测年限的变化基本趋势和封育样地内类似,但物种数最多年份却是开始封育的2003年,说明连续放牧降低了样地内的物种数。封育对高寒草甸样地内物种数的影响主要表现为封育后期,封育6年时样地内的物种数显著小于未封育样地内的物种数。
2.2 封育对物种多样性的影响
监测结果表明,持续围栏封育对高寒草甸群落植物物种多样性具有明显的影响。封育和未封育高寒草甸群落内植物丰富度指数随着封育年限变化趋势基本类似,均表现为先显著下降,后显著增加,再显著下降(P<0.05),但封育群落内植物丰富度指数最高出现在封育4年时,而未封育群落内植物丰富度指数最大值却出现在封育第1年(图1)。群落内均匀度指数变化在封育和未封育条件下出现明显的分异,封育群落内均匀度指数表现为先显著下降,后显著增加,再显著下降(P<0.05),其最大值出现在封育4年时,而未封育群落内均匀度指数却表现为先显著增加,后显著降低(P<0.05),其最大值出现在第3年和第5年(图1)。多样性指数综合反映了群落内物种丰富程度和种群分布均匀程度,封育群落的植物多样性指数随着封育年限的变化趋势趋同于均匀度指数和丰富度指数的变化趋势,而未封育群落植物多样性指数随着监测年限的变化趋势趋同于均匀度指数变化趋势,而趋异于丰富度指数的变化趋势(图1)。封育时群落内多样性指数与丰富度指数的相关方程为y=11.37x+3.575(R=0.913,P<0.05),而与均匀度指数的相关方程为y=0.223x+0.163 6(R=0.981,P<0.05),说明此时多样性指数与均匀度指数的相关性大于其与丰富度指数的关系;未封育群落内多样性指数与丰富度指数相关系数仅为0.225,且不显著,而多样性指数与均匀度指数的相关方程为y=0.331 9x-0.023 4(R=0.967,P<0.05)。

表1 封育对高寒草甸物种数及其所属的属数和科数的影响Table 1 Effect of fence on plant species,number of genera and families in the alpine meadow
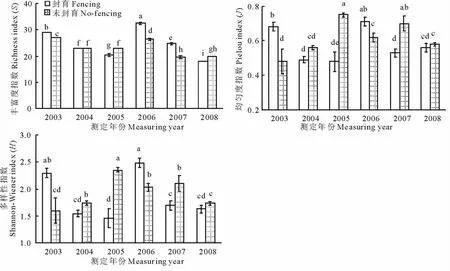
图1 封育对高寒草甸群落植物丰富度指数,均匀度指数和多样性指数的影响Fig.1 Effect of fence on richness index,evenness index and diversity index of alpine meadow communities
2.3 封育对群落高度的影响
未封育群落和封育群落高度随着监测年限的变化趋势类似,均表现为先显著下降,后显著增高(P<0.05),封育群落高度最大值出现在封育第5年和第6年(图2),未封育群落高度最大值出现于监测第6年。封育群落高度和未封育群落高度间的关系随着封育年限延长而发生改变,封育第1年虽然封育群落高度略大于未封育群落,但两者差异并不显著,第2年和第3年均表现为封育群落高度显著小于未封育群落(P<0.05),从第4年开始封育群落高度显著大于未封育群落(P<0.05),说明封育和未封育群落高度转换发生于封育第4年。
2.4 封育对群落盖度的影响
未封育和封育群落盖度随着封育年限延长均表现为先显著下降,后显著上升,再显著下降的变化趋势(P<0.05),两者最大盖度均出现在封育第4年(图2)。封育群落盖度和未封育间的关系随着封育时间延长发生明显的变化,封育第1年,两者盖度差异不大,第2年和第3年封育群落盖度显著小于未封育群落(P<0.05),到封育第4年两者的差异不显著,自第5年开始,封育群落盖度显著大于未封育群落(P<0.05),说明封育年限对群落盖度的影响在第4年发生转变(图2)。

图2 封育对高寒草甸群落高度和盖度的影响Fig.2 Effect of fence on height and cover of alpine meadow communities
2.5 封育对群落地上生物量的影响
封育草甸和未封育草甸生物量随着监测年限延长出现明显的分异现象。未封育草甸生物量随着监测年限延长开始时逐渐下降,但下降幅度不显著,而自第5年下降幅度达到显著水平(P<0.05)。封育草甸生物量随着封育时间延长表现为先显著增加后显著降低(P<0.05),封育第4年和第5年草甸生物量最大,这说明封育初期促进了高寒草甸初级生产力的提高,但超过一定的封育年限后,高寒草甸初级生产力开始下降。封育和未封育草甸生物量间的关系随着封育年限延长发生一定的变化,封育前3年内,两者间的差异不显著,而自第4年开始封育草甸生物量显著大于未封育(P<0.05)(图3)。
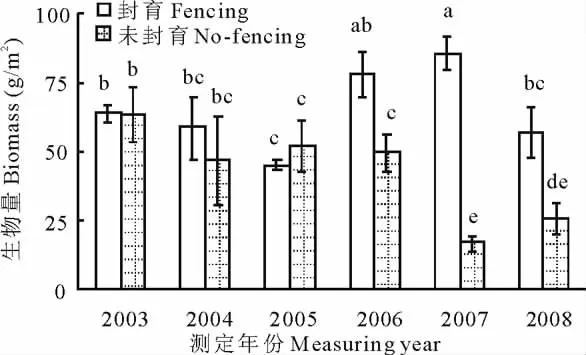
图3 封育对高寒草甸生物量的影响Fig.3 Effect of fence on biomass of alpine meadow
3 讨论
封育是人为干扰草地的一种方式,其实质是采用围栏手段避免草地受大型食草动物对草地的采食和践踏,达到保护或者恢复退化草地的目标,但客观上部分切割了自然草地生态系统食物链和食物网,因此,其对植物群落具有积极和消极双重作用[17],而草地管理的目标就是扩展其积极作用,抑制或者削弱其消极作用。草地封育后由于缺少大型食草动物的采食,家畜喜食植物受抑制程度减弱,生长加快,较放牧草地而言对家畜不喜食植物竞争力加强,使群落组分、高度、盖度均发生一定的变化,引发物种多样性和生物量发生变化。研究结果表明,围栏提高了新疆天山地区草地的生物量、高度、盖度和频度[18],但封育对草地群落物种多样性的影响出现分异,表现为封育降低了新疆天山地区[18]和河北北部坝上地区[19]草地群落的物种多样性,提高了尼泊尔特莱地区[20]和澳大利亚东南部地区[21]草原群落物种多样性,但对澳大利亚南部地区的草地群落植物多样性没有显著影响[22],这些分异产生的原因要么是封育时间不一致所致,要么是草地生境或类型不一样所致。本研究结果表明,封育初期高寒草甸群落内物种数、高度、盖度和物种多样性均呈下降趋势,至封育第4年开始上升,第5或第6年又开始下降,表明封育对高寒草甸群落特征的影响呈单峰现象,这种变化趋势和内蒙古草原长期围栏封育过程中群落多样性的变化趋势一致[23],但2个地区植物多样性发生转变的封育年限不同,内蒙古草原群落多样性对封育响应的拐点是12年[23],典型草原的适宜围栏封育年限为14年[24],而天祝高寒草甸群落物种多样性对封育的响应拐点是4年,主要原因是内蒙古草原降水量少,群落植物生长受降水影响较相对湿润的天祝高寒草甸大。
Willms等[25]对加拿大阿尔伯达省北部大平原草地的研究表明,围封封育对草地初级生产力影响不大,而围栏降低了内蒙古羊草(Leymuschinensis)草原草地初级生产力,中度放牧提高了草地初级生产力[26]。本研究结果表明,6年围栏封育时期内,高寒草甸初级生产力先降低后增至最大值,然后又下降,说明围栏封育对草地初级生产力的影响不仅与草地类型有关,而且与草地分布的生境有关,也可能与封育期的气候环境有关。
因此,综合考虑草地群落的组分、高度、盖度、物种多样性和初级生产力,位于青藏高原东北边缘的甘肃天祝高寒草甸适宜围栏封育年限为4年。
[1]张法伟,李跃清,李英年.高寒草甸不同功能群植被盖度对模拟气候变化的短期响应[J].草业学报,2010,19(6):72-78.
[2]Guo Z G,Long R J,Niu F J,etal.Effect of highway construction on plant diversity of grassland communities in the permafrost regions of the Qinghai-Tibet plateau[J].The Rangeland Journal,2007,29:161-167.
[3]郭正刚,牛富俊,湛虎,等.青藏高原北部多年冻土退化过程中生态系统的变化特征[J].生态学报,2007,27(8):3294-3301.
[4]包维楷,张镱锂,摆万奇.青藏高原东部采伐迹地早期人工重建序列梯度上植物多样性的变化[J].植物生态学报,2002,26(3):330-338.
[5]韩龙,郭彦军,韩建国.不同刈割强度下羊草草甸草原生物量与植物群落多样性研究[J].草业学报,2010,19(3):70-75.
[6]焦婷,常根柱,周学辉.高寒草甸草场不同载畜量下土壤酶与土壤肥力的关系研究[J].草业学报,2009,18(6):98-104.
[7]Gonzales E K,Clements D R.Plant community biomass shifts in response to mowing and fencing in invaded oak meadows with non-native grasses and abundant ungulates restoration[J].Ecology,2010,18(5):753-761.
[8]王英舜,师桂花,许中旗,等.锡林郭勒放牧草地封育后植被恢复过程的研究[J].草业科学,2010,27(8):10-14.
[9]范燕敏,孙宗玖,武红旗,等.封育对山地草地植被及土壤特性的影响[J].草业科学,2009,26(3):79-82.
[10]刘丙万,蒋志刚.青海湖草原围栏对植物群落的影响兼论濒危动物普氏原羚的保护[J].生物多样性,2002,10(3):326-331.
[11]吴宁,罗鹏.长江上游高寒草地生态建设和管理中生态理论的若干质疑[J].应用与环境生物学报,2004,10(4):537-542.
[12]侯扶江,任继周.甘肃马鹿冬季放牧践踏作用及其对土壤理化性质影响的评价[J].生态学报,2003,23(3):486-495.
[13]杨成德,陈秀蓉,龙瑞军.东祁连山高寒草地返青期不同草地型土壤氮的分布特征[J].草业学报,2010,19(1):67-74.
[14]Wang H,Guo Z G,Xu X H,etal.Response of vegetation and soils to desertification of alpine meadow in the upper basin of the Yellow River,China[J].New Zealand Journal of Agricultural Research,2007,50:491-501.
[15]宋理明,马晓虹,魏永林.海北州天然草地退化沙化成因与对策[J].草业科学,2010,26(7):186-190.
[16]Grieg S P.Quantitative Plant Ecology[M].Oxford:Blackwell Scientific Publications,1983.
[17]江小蕾,张卫国,杨振宇.不同干扰类型对高寒草甸群落结构和植物多样性的影响[J].西北植物学报,2003,23(9):1479-1485.
[18]李慧,蒋平安,程路明.围栏对新疆山区草地植物群落多样性的影响[J].新疆农业大学学报,2005,28(3):40-43.
[19]张洪生,邵新庆,刘贵河.围封、浅耕翻改良技术对退化羊草草地植被恢复的影响[J].草地学报,2010,18(3):339-344.
[20]Karki J B,Yadavendardev V.Grazing lawns in Terai grasslands,Royal Bardia National Park,Nepal[J].Biotropica,2000,32(3):423-429.
[21]Briggs S V,Taws N M.Condition of fenced and unfenced remnant vegetation in inland catchments in south-eastern Australia[J].Australian Journal of Botany,2008,56(7):590-599.
[22]Meissner R A,Facelli J M.Effect of sheep exclusion on the soil seed bank and annual vegetation in chenopod shrub lands of South Australia[J].Journal of Arid Environment,1999,42:117-128.
[23]宝音陶格涛,刘海松,图雅.退化羊草草原围栏封育多样性动态研究[J].中国草地学报,2009,31(5):37-40.
[24]单贵莲,徐柱.围封年限对典型草原群落结构及物种多样性的影响[J].草业学报,2008,17(6):1-8.
[25]Willms W D,Smoliak S,Dormaar J F.Vegetation response to time-controlled grazing on mixed and fescue prairie[J].Journal of Range Management,1990,43(6):513-517.
[26]王明君,韩国栋,崔国文,等.放牧强度对草甸草原生产力和多样性的影响[J].生态学杂志,2010,29(5):862-868.
