不同游泳运动方式对大鼠海马cAMP影响的实验研究
2011-12-28张安民张瑞萍贾慧娟王鹏程
张安民,张瑞萍,贾慧娟,王鹏程
(1.烟台大学 体育学院,山东 烟台 264005;2.山西大学 体育学院,太原 030006)
不同游泳运动方式对大鼠海马cAMP影响的实验研究
张安民1,张瑞萍1,贾慧娟2,王鹏程1
(1.烟台大学 体育学院,山东 烟台 264005;2.山西大学 体育学院,太原 030006)
目的:观察游泳运动后大鼠海马cAMP浓度变化,探讨不同游泳运动方式对海马cAMP的影响机制。方法:建立大鼠游泳运动模型 (持续运动、间歇运动),采用酶联接免疫吸附剂法,观察大鼠海马cAMP浓度变化,并用统计学软件进行数据分析。结果:运动后持续组和间歇组cAMP浓度均呈锯齿形特征,但变化规律有明显差异。持续组先降后升,与对照组相比在即刻、60min、120min、240min有所下降,有显著差异 (p<0.05);在60min达到最低值 (p<0.01);运动后30min略高于对照组,无显著差异性 (p>0.05)。间歇组先升后降且变化幅度逐渐增大,与对照组相比运动后240min cAMP浓度达到峰值,有显著性差异 (p<0.01);而运动后30min、120min cAMP浓度呈下降趋势,有显著性差异 (p<0.01);运动后即刻、60min cAMP浓度略有上升,无显著差异 (p>0.05)。结论:不同游泳运动方式对海马cAMP的影响不同,具有一定的时效性。
海马;第二信使cAMP;持续游泳运动;间歇游泳运动
运动能促进人的认知功能和个性发展,特别是学习记忆这种脑功能的高级形式[1]。大量动物实验也表明,运动有助于动物海马等与学习记忆密切相关脑区神经元的增殖、存活和分化,促进长时程增强的诱导,提高实验动物的空间记忆和被动逃避记忆能力[2-3]。cAMP是最早确定的细胞内的第二信使,作为细胞功能的信号传导物质参与学习记忆过程[4-5]。因此,研究海马与cAMP的表达变化将会对海马的学习记忆过程有重要意义。本实验旨在探讨不同游泳运动方式对大鼠海马cAMP表达的时间变化规律及其可能的影响机制。
1 材料和方法
1.1 实验动物和分组
实验动物:SD成年雄性大鼠55只,鼠龄3个月,体重250-300g,由山西医科大学动物实验中心提供。饲养条件:分笼饲养,每笼4只,自由饮食,自然昼夜节律光照,室温20±5℃。相对湿度在40%至60%。将大鼠随机分为对照组 (n=5只)、持续运动组 (n=25只)、间歇运动组(n=25只)。
1.2 运动模型建立
采用游泳运动方式建立运动动物模型。游泳条件:塑钢玻璃游泳池,150cm×60cm×70cm,水深为大鼠身体长度的2倍,约60cm,水温保持在33℃ 至36℃ 之间,每个游泳池随机分配5只大鼠。正式训练前运动组大鼠进行3天适应性游泳训练,三天的运动时间每天分别为15min、20 min、30min。随后开始正式的游泳运动训练,每周训练6次,共训练6周。负重为体重5%,游泳6min,休息4 min,每天连续进行10组。对照组不做任何运动,常规饲养,自由活动。训练结束后吹干毛发,返回鼠笼。
1.3 体重测量
采用HD-2006B天平,对大鼠的体重进行全程测量。开始运动前测量一次,然后每周一运动前对所有大鼠进行体重测量、记录,便于统计分析。
1.4 取材
最后一次游泳后,持续组和间歇组大鼠按运动后即刻、30min、60min、120min、240min五个时刻分别依次取材,操作如下:大鼠腹腔2%戊巴比妥钠麻醉 (40mg/kg),断头,冰上取脑,置于-80℃液氮冷藏。对照组操作同上。
从液氮取出大鼠脑组织,复温至常温。根据大鼠立体定向图谱,分离脑组织切取大鼠海马组织约50mg—100mg,置于离心管,按10mg∶200ul比例加入0.01MPBS液,用超声波细胞粉碎机将其制成乳白色悬液,12000r/min离心10分钟,取上清液放入-4℃冰箱保存备用。
1.5 cAMP含量的测定
cAMP浓度的测定均采用双抗体酶联免疫吸附法 (Enzymes linked immunosorbant assays,ELISA法),大鼠环磷腺苷cAMP ELISA试剂盒购自美国R·B公司,均严格按说明书进行操作。检测仪器采用Benchmark酶标仪 (BIORAD公司)。
具体操作步骤:①加样:将100ul标准品、100ul标本加入相应反应板孔中;②轻轻混匀30s,封住板孔,37℃温育90min;③洗板:甩尽板内液体,用洗涤液洗涤反应板 (每孔内加入350ul洗涤液),并去除水滴 (在厚叠吸水纸上拍干);反复洗涤5次;④每孔加入100ul 1×Biotin。轻轻混匀30s,封住板孔,37℃温育60min;⑤洗板:甩尽板内液体,用洗涤液洗涤反应板 (每孔内加入350ul洗涤液),并去除水滴 (在厚叠吸水纸上拍干);反复洗涤5次;⑥每孔加入100ul 1×HRP。轻轻混匀30s,封住板孔,37℃温育30min;⑦洗板:甩尽板内液体,用洗涤液洗涤反应板 (每孔内加入350ul洗涤液),并去除水滴 (在厚叠吸水纸上拍干);反复洗涤5次;⑧每孔加入50ul显色剂A和50ul显色液B,轻轻混匀10s,37℃暗处温育15±10min;⑨每孔加入100ul终止液 (Stop Solu-tion)。轻轻混匀30s;30min内在450nm处读OD值;⑩以OD值为纵坐标,以标准品浓度为横坐标,绘制标准曲线。并根据样本的OD值查出其浓度,或用回归方程计算其浓度。
1.6 统计学方法
采用SPSS12.0软件对实验数据进行统计处理。所有数据均用平均数±s标准差表示,组间比较采用t检验,p<0.05有统计学意义。
2 实验结果
2.1 不同游泳运动方式对大鼠体重的影响
从表1可以看出,不同游泳运动方式对大鼠体重影响非常明显。训练前三组大鼠 (对照组、持续组、间歇组)体重相比无显著差异 (P>0.05);训练后对照组大鼠体重增加较快;持续组大鼠体重在第2周明显下降,随后保持稳定;间歇组大鼠体重略有增长,但组间无显著性差异。间歇组、持续组大鼠体重均低于对照组,且具有非常显著性差异 (P<0.01)。说明两种游泳训练都有一定的运动强度,对大鼠的控体重有一定的影响。

表1 游泳训练后大鼠体重影响结果
2.2 不同游泳运动方式对大鼠海马cAMP浓度的影响
2.2.1 持续游泳运动对大鼠海马cAMP浓度的影响
统计学处理结果显示 (表2):持续游泳运动后cAMP浓度先降后升,与对照组相比在即刻、60min、120min、240min有所下降,有显著差异 (p<0.05);在60min达到最低值,有非常显著性差异 (p<0.01);运动后30min略高于对照组,无显著差异性 (p>0.05)。
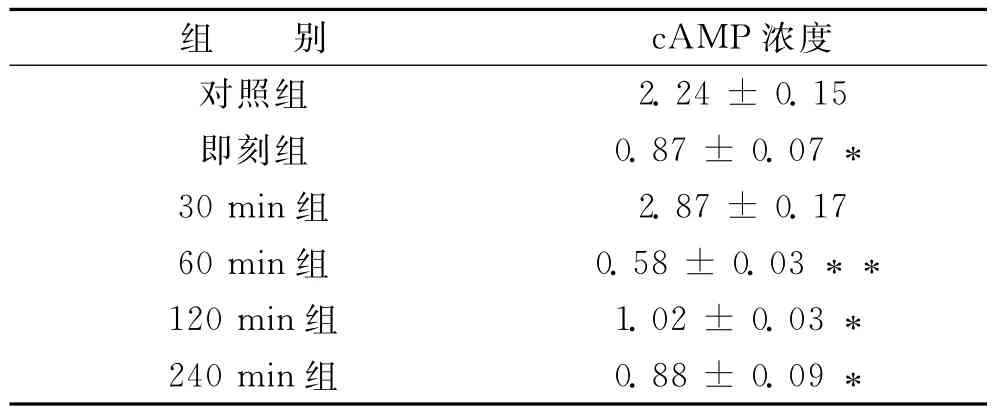
表2 大鼠海马在五个不同时刻cAMP浓度变化 (持续游泳)
从图1来看,运动后海马cAMP浓度随时间呈锯齿形的变化,30min组上升幅度较大,随后快速下降,120min、240min变化幅度逐渐趋于平缓。
2.2.2 间歇游泳运动对大鼠海马cAMP浓度的影响
统计学处理结果显示 (表3):间歇游泳运动后cAMP浓度先升后降且变化幅度逐渐增大,与对照组相比运动后240min cAMP浓度达到峰值,有显著性差异 (p<0.01);而运动后30min、120min cAMP浓度呈下降趋势,有显著性差异 (p<0.01);运动后即刻、60min cAMP浓度略有上升,无显著差异 (p>0.05)。
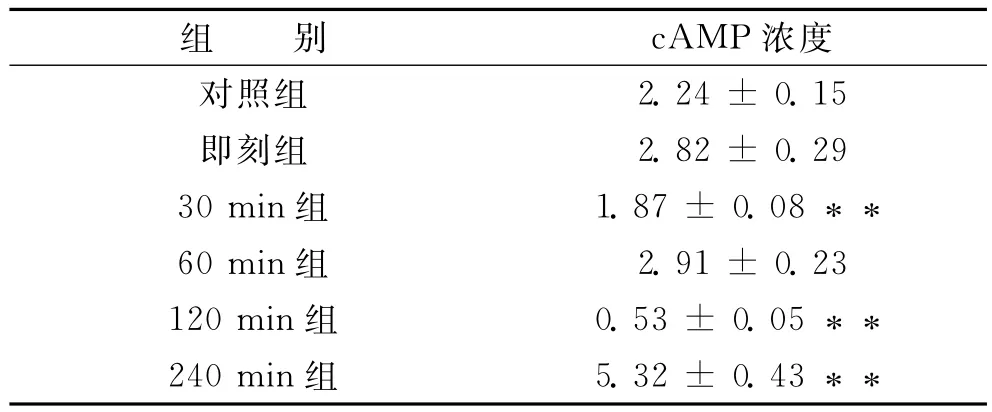
表3 大鼠海马在五个不同时刻cAMP浓度变化 (间歇游泳)
从图2来看,运动后,海马cAMP浓度随时间呈锯齿形的变化,即刻、60min组变化幅度较小,随后变化幅度逐渐增大,240min cAMP浓度突然升高,达到峰值。

图1 持续游泳运动对大鼠海马cAMP在不同时刻表达的时效性

图2 间歇游泳运动对大鼠海马cAMP在不同时刻表达的时效性
3 讨论与分析
经过6周的游泳运动训练:持续运动组大鼠体重略有下降但基本保持稳定;间歇运动组大鼠体重增加相对缓慢,皮毛光亮,体形匀称。对照组大鼠体重增加较快,显得臃肿。说明两种游泳训练都有一定的运动强度,对大鼠的控体重可能有一定的影响,证明游泳运动模型的建立较成功。
cAMP是细胞内第二信使,参与学习和记忆过程[4-5]。在生理状态下,神经细胞中cAMP由三磷酸腺苷 (ATP)经腺苷环化酶 (AC)催化生成,由磷酸二酯酶 (PDE)降解,二者的功能状态维持神经细胞cAMP水平稳定。第二信使的特点是在短时间内即可迅速产生,短时间内又可迅速灭活,从而具有信号传递的功能。也就是说,在外来信号的作用下,第二信使上游的信号传导蛋白被激活后,利用特殊的原料,在短时间内催化生成大量的第二信使,实现从胞外信号到胞内的转换和放大,同时在特定机制作用下,生成的大量第二信使被迅速降解或转移,使细胞内第二信使的浓度迅速降低,以保证信号转导的一过性。
运动作为典型的应激源,对cAMP浓度具有显著性影响。罗艳蕊等[6]研究表明8周游泳训练导致心脏代偿性肥大,cAMP/cGMP比值减小。黄元汛、张钧[7]研究发现,力竭运动使大鼠心肌中cAMP和cGMP均明显高于对照组,说明力竭运动可造成心肌中cAMP和cGMP含量明显升高。张安民[8-10]等研究发现持续游泳训练和间歇性负重游泳训练能够引起纹状体cAMP浓度升高并产生后续时程变化,并且认为这种变化可能与c-fos原癌基因、神经细胞内钙离子等有关。Zimmer[11]对实验性压力超负荷动物模型的研究发现,压力超负荷与心肌cAMP和心肌肥厚显著正相关;压力超负荷诱导了心肌cAMP浓度上升,同时cGMP浓度下降,cAMP/cGMP比值增大,推测环核苷酸系统 (cAMP/cGMP)可能介导了压力超负荷导致心肌肥大的反应。同时心肌组织cAMP含量于术后0.5h明显增加,术后5d时达峰值,术后30d降至正常。李卫平等[12]实验表明:服用抗氧化剂对耐力运动组和间歇运动组恢复期的心肌cAMP浓度具有一定的影响,而对这两组恢复期的心肌cGMP浓度则并无显著性作用;运动方式和抗氧化剂是影响心肌cAMP、cGMP浓度变化的主要因素。徐松德等[13]将34只家兔随机分为对照、运动和不运动三组,后两组每只家兔喂胆固醇200mg/d,试验四周后结果表明运动组动脉壁cAMP水平增加19.2%,不运动组则降低49.4%;肝脏cAMP含量运动组与不运动组分别降低21.2%和15.7%,血浆cAMP浓度运动组与不运动组分别升高0.95%和6.8%。王泽军等[14]研究表明游泳训练可以使大鼠海马cAMP浓度分别发生了显著的升高。Bernabeu[15]在实验中发现大鼠海马cGMP水平在训练后即刻达峰值,相反cAMP水平在3h后才达峰值。
本实验把间歇运动和持续运动作为应激源对大鼠进行研究,该二者是两种完全不同的运动:前者是一种极限强度并且严格控制运动和间歇时间的无氧训练方法[16],后者具有强度低、有节奏、持续时间较长的特点。通过六周的游泳训练,结果显示,不同游泳运动对大鼠海马cAMP影响不同。二者虽然都呈锯齿形特征,但变化规律相差甚远。间歇运动组先升后降且幅度逐渐增大,运动后即刻与对照组相比略有上升,但无显著差异 (p>0.05),运动后30min cAMP浓度呈下降趋势,有显著性差异 (p<0.01);60min cAMP浓度略有上升,无显著差异 (p>0.05),运动后120min cAMP浓度呈下降趋势,有显著性差异 (p<0.01);运动后240min cAMP浓度达到峰值,有显著性差异 (p<0.01)。原因可能是由于长期的运动训练刺激,使cAMP一PKA通路中的某些激素受体的适应能力增强所致。在cAMP/PKACREB信号通路中,活化的PKA催化亚基可以通过细胞内的转位而接近底物蛋白CREB,使CREB磷酸化,调节转录因子的活性[17],从而介导细胞对外界刺激 (如运动)的反应。而CREB作为核内 “第三信使”,被认为是对外部刺激产生适应以及长时记忆的最适合转录因子。磷酸化的CREB可以激活相关基因的转录,其中包括调节C-FOS以及通过Ca2+依赖性的形式调节 BDNF的表达[18-19]。间歇组cAMP浓度在240min达到峰值说明在这一时刻可能海马的学习记忆能力得到了很大的提高。持续运动组先降后升,与对照组相比在即刻有所下降,有显著差异 (p<0.05);运动后30 min略高于对照组,但无显著差异性 (p>0.05);在60min达到最低值 (p<0.01);120min有所上升但仍低于对照组,240min略有下降,二者均有显著差异 (p<0.05)。持续组cAMP浓度只有在运动后30min略高于对照组且无明显差异,其他时刻均低于对照组。其变化比较平缓可能和对运动的适应、cAMP的一过性等特点有关。有研究发现cRE BmRNA升高是在运动结束后第3d以及第7d,在第28d cREBmRNA水平并无显著变化[20]。总之,不同运动方式(运动强度、运动时间、运动间歇等)对海马cAMP浓度变化存在一定的时效性 (表达峰值、表达持续时间等)。
海马cAMP浓度的变化和运动强度、运动时间、运动量有很大的关系[21]。疾病、运动、药物等多种刺激因素均可引起神经细胞内cAMP浓度发生改变继而引发后续各种反应[22-32]。研究不同运动方式对大鼠海马cAMP浓度的影响,把体育锻炼作为一种积极而有效的方法,可以为在学习记忆或脑功能方面有缺陷的患者提供一定的帮助,同时可以为制定运动处方提供实验依据。细胞信号传导过程极为复杂,多是通过多条信息转导的分支与汇聚来实现的。cAMP的变化又是如何影响PKA、CREB等相关蛋白激酶和转录因子的,这些问题的提出与解决,有助于我们今后更深入的研究。究竟何种方式、强度及持续时间的运动对学习记忆的提高最适宜?这将是继续研究的关键问题。
[1] Uysal N,Tugyan K,Kayatekin B M,et al.The effects of regular aerobic exercise in adolescent period on hippocampal neurondensity,apoptosis and spatial memory[J].Neuroscience Letters,2005,383(3):241-245.
[2] Radak Z,Kaneko T,Tahara S,et al.Regular exercise improves cognitive function and decreases oxidative damage in rat brain [J].Neurochemistry Int,2001,38(1):17-23.
[3] Adlard P A,Perreau V M,Cesar C E,et al.The Timecourse of induction of brain-derived neurotrophic factor mRNA and protein in the rat hippocampus following voluntary exercise[J].Neuroscience Letters,2004,363(1):43-48.
[4] Nazif F A ,Byren J H,Cleary L J.cAMP induces long-term morphological changes in sensory neurons of Aplysia[J].Brain Research,1991(539)∶324-327.
[5] Vianna M R,Izquierdo L A,Barros D M,et al.Differentialrole of hippocampal cAMP -dependent protein kinase inshort and long-term memory[J].Neurochemistry Research,2000,25(5):621-626.
[6] 罗艳蕊,漆正堂,贺杰.游泳训练后大鼠心肌细胞cAMP、cGMP含量的变化[J].中国应用生理学杂志,2005,21(2):185-186.
[7] 黄元汛,张钧.牛磺酸对大鼠力竭运动时第二信使及心肌线粒体中钙离子含量变化的影响[J].中国运动医学杂志,1999,18(2):130-133.
[8] 张安民,张青元,于芳,等.不同方式游泳训练对大鼠纹状体cAMP浓度影响的实验研究[J].体育科学,2010,30(1):62-65.
[9] 张安民,王根深,于芳,等.大鼠持续运动对中央杏仁核c-fos蛋白表达影响的实验研究[J].体育科学,2008,28(1):39-42.
[10] 张安民,张青元,于芳,等.持续游泳运动对大鼠脑杏仁核神经细胞内钙离子的影响[J].成都体育学院学报,2009,(35)6:70-72.
[11] Zimmer H G,Petter H.Metabolic aspects of the development of experimental cardiac hypertrophy[J].Basic Res Cardiol,1986(81):127-137.
[12] 李卫平,陈峰,吴正平.不同运动方式和抗氧化剂对大鼠心肌cAMP、cGMP代谢的影响[J].武汉体育学院学报,2008,42(5):85-87.
[13] 徐松德,吴湘云,刘贤锡,等.运动对喂胆固醇家兔血浆动脉壁及肝脏水平的影响[J].山东医学院学报,1985,23(2):40-43.
[14] 王泽军.游泳训练对大鼠空间学习记忆力和脑内NO/·OH-cGMP、cAMP的影响[D].上海:上海华东师范大学,2007.
[15] Bernabeu R,Bevilaqua L,Ardenghi P,et al.Involvement of hippocampal cAMP/cAMP-dependent protein kinase signaling pathways in a late memory consolidation phase of aversively motivated Learning in rats[J].Proc Natl Acad Sci USA,1998(94):7041-7046.
[16] 洪长青,邓树勋.三种训练对大鼠心肌细胞超微结构的影响[J].浙江体育学,2000,22(3):55-58.
[17] 李俊发,贺俊崎.细胞信号转导研究技术[M].北京:中国协和医科大学出版社,2008:32.
[18] 于芳,张安民,王根深,等.间歇性负重游泳训练对大鼠杏仁核基底外侧核(BLA)C-FOS蛋白表达的影响[J].北京体育大学学报,2008,31(10):1357-1360.
[19] 罗德生.钙调蛋白与cAMP代谢[J].临床荟萃,1996,11(9):421-422.
[20] Molteni R,Ying Z,Mezpinilla Fgo′.Differential effects of acute and chronic exercise on plasticity-related genes in the rat hippocampus revealed by microarray[J].Eur Neurosci,2002(16):1107-1116.
[21] 潘华山,赖秋媛,冯毅翀,等.运动氧化应激机制与人参皂甙Rb1抗运动性疲劳的实验研究[J].河北体育学院学报,2010,24(2):68-70.
[22] 刘晶,席宏杰,王淑云,等.MK-801对吗啡依赖大鼠戒断症状及中枢cAMP含量的影响[J].中国药理学通报,2006,22(9):1115-1118.
[23] 杨亚安,邵君飞,陈尔齐,等.小鼠海马组织中cAMP含量和AC活性的昼夜变化[J].苏州大学学报,2003,23(1):8-9.
[24] 周爽,孙国杰,杨毅红.针刺对急性脑梗塞患者血浆中cAMP、cGMP含量的影响[J].针灸临床杂志,2000,16(4):51-52.
[25] 汤燕平,白若华.高原低氧对鼠脑cAMP和cGMP含量的影响[J].青海医学院学报,1993,35(14):165-166.
[26] 刘洁,赵胜利,姬汴生.人参对衰老大鼠Β受体-cAMP系统的影响及其机理初探[J].中药药理与临床,1995(6):22-24.
[27] 朱金华.Υ辐照对小鼠中枢核团细胞中cAMP、cGMP昼夜节律的影响[J].江西预防医学,2001,12(2):7-9.
[28] 王小艳,董军,陆大祥.气味对小鼠学习记忆能力及海马cAMP反应元件结合蛋白的影响[J].中国病理生理杂志,2004,20(3):367-370.
[29] 曲润江,郭鹤,郑振兴.电磁波脉冲对大鼠松果体及血浆中cAMP和cGMP的影响[J].第四军医大学学报,2000,21(11):1314-1316.
[30] 王伦安.cAMP在疾病过程中的作用[J].华西医学,2003,18(2):293-294.
[31] 杨亚安,邵君飞,陈尔齐,等.小鼠海马组织中cAMP含量和AC活性的昼夜变化[J].苏州大学学报,2003,23(1):8-9.
[32] 张云东,朱佩芳,王正国,等.CRI-I对培养下丘脑神经元胞浆内cAMP和Ca2+变化的影响[J].第三军医大学学报,2003,25(12):1059-1061.
The Experimental Study on the Effect of Different Swimming Ways on the Rat Hippocampus’s cAMP
ZHANG An-min1,ZHANG Rui-ping1,JIA Hui-juan2,WANG Peng-cheng1
(1.College of Physical Education,Yantai University,Yantai 264005,China;2.College of Physical Education,Shanxi University,Taiyuan 030006,China)
Objective:To observe the change of cAMP concentration in rat hippocampus after swimming and to explore different swimming ways’effect on the hippocampal cAMP.Methods:To establish a rat swimming model(continued swimming,intermittent swimming),using enzyme link immunosorbent method to observe the changes of hippocampal cAMP concentration,and use statistical software for data analysis.Results:After exercise the continued group and the intermittent group’s cAMP concentration all appeared the shape of sawteeth,but there are significant differences in variation.Continued group falls first and then rises,compared with the control group,it declines in the immediate,60min,120min,240min,there are significant differences(p<0.05);in 60min it reaches the minimum (p<0.01);slightly higher than the control group 30min after exercise,there was no significant difference(p>0.05).The intermittent group increases first and then decreases gradually and the magnitude changes,as compared with the control group,the cAMP concentration reaches its peak 240min after exercise,there is a significant difference(P<0.01);and 30min,120min after exercise,cAMP concentration is in a downward trend,there was a significant difference(P<0.01);The cAMP concentration increases slightly immediately after exercise,60min,there is no significant difference(p>0.05).Conclusion:Different ways of swimming have different impact on hippocampal cAMP and it has a certain timeliness.
hippocampus;second messenger cAMP;endurance swimming;intermittent swimming
G804.7
A
1008-3596(2011)06-0078-05
2011-08-10
国家自然科学
(30671019);山东省自然科学基金项目(Y2008c29)
张安民 (1959-),男,山西万荣人,教授,研究方向为运动医学。
