牛角藓Cratoneuronfilicinum(Hedw.)Spruce的RAPD分类研究
2011-12-22李鹏勃吴玉环
李鹏勃,吴玉环,*,罗 昊
(1.杭州师范大学生命与环境科学学院,浙江杭州 310036;2.中国科学院沈阳应用生态研究所,辽宁沈阳 110016)
牛角藓Cratoneuronfilicinum(Hedw.)Spruce的RAPD分类研究
李鹏勃1,吴玉环1,2*,罗 昊2
(1.杭州师范大学生命与环境科学学院,浙江杭州 310036;2.中国科学院沈阳应用生态研究所,辽宁沈阳 110016)
以采自四川、甘肃、湖南等8省的9份牛角藓标本为材料,运用RAPD技术对牛角藓的分类修订进行研究.利用15条随机引物共获得134条清晰谱带,其中多态性条带占91.79%,牛角藓各标本间的遗传相似系数为0.703~0.870.结果显示:虽然9份牛角藓标本的形态特征差异非常大,但通过RAPD分析表明其为同一种.
牛角藓;RAPD;分类修订;环境饰变
牛角藓属(Cratoneuron(Sull.)Spruce)是薄罗藓亚目(Leskeineae)柳叶藓科(Amblystegiaceae)的一个属,现记录仅有1种1变种[1],即牛角藓(Cratoneuronfilicinum(Hedw.)Spruce)和牛角藓宽肋变种(Cratoneuronfilicinum(Hedw.)Spruce var.atrovirens(Brid.)Ochyra),广泛分布于日本、俄罗斯,东南亚、欧洲、北美和北非等地,我国东北、秦岭沿线、西南地区及新疆等地也有较为丰富的资源[2].
牛角藓植物体大小、叶形和角部细胞等形态特征变异非常大,且表现出明显的依生态环境(尤其是固着基质和水湿条件)不同而相异的特点[3],因此对经典形态分类学结果造成了较大的影响,以致以前的学者们根据是否具鳞毛、中肋的粗细、叶形和角部细胞的不同变化等将大量牛角藓标本作为新种发表,其中以中国牛角藓植物标本发表的新种就有17个[4-6].而Ochyra[1]则认为全世界牛角藓属仅存牛角藓及其宽肋变种,吴玉环等[7]对200余份牛角藓标本的叶形、中肋和角部细胞等特征数据进行线性回归分析也得到了相似的结果.因此,急需寻求一种新的、更加稳定可靠的研究方法来确定这些叶形和中肋等形态特征变化较大的牛角藓是否可以简单地归于一种.
RAPD技术因其简单、快速、高效等优点,加之在苔藓植物研究中应用时,能够避免自身不能区分纯合和杂合等劣势[8],近年来已在苔藓植物遗传多样性检测、遗传关系讨论、遗传图谱构建、基因定位与分离等方面得到了广泛的应用[9-11].
该文拟通过分子生物学手段对采自四川、甘肃、湖南等8个省份9个居群的牛角藓进行RAPD分类研究,探讨中国牛角藓不同居群的遗传变异程度,并结合近年来经典形态分类学所取得的研究成果,提出关于中国牛角藓分类的新观点,从期能为经典形态分类学的研究成果提供新的分子实验证据,并为其他科属苔藓植物的系统分类修订提供新的借鉴和参考.
1 材料与方法
1.1 材 料
9份采于不同地区及不同生态环境的牛角藓标本(表1),选取牛角藓宽肋变种和柳叶藓(Amblystegiumserpens)作为外类群.所用标本均由沈阳应用生态研究所标本馆(IFSBH)提供.
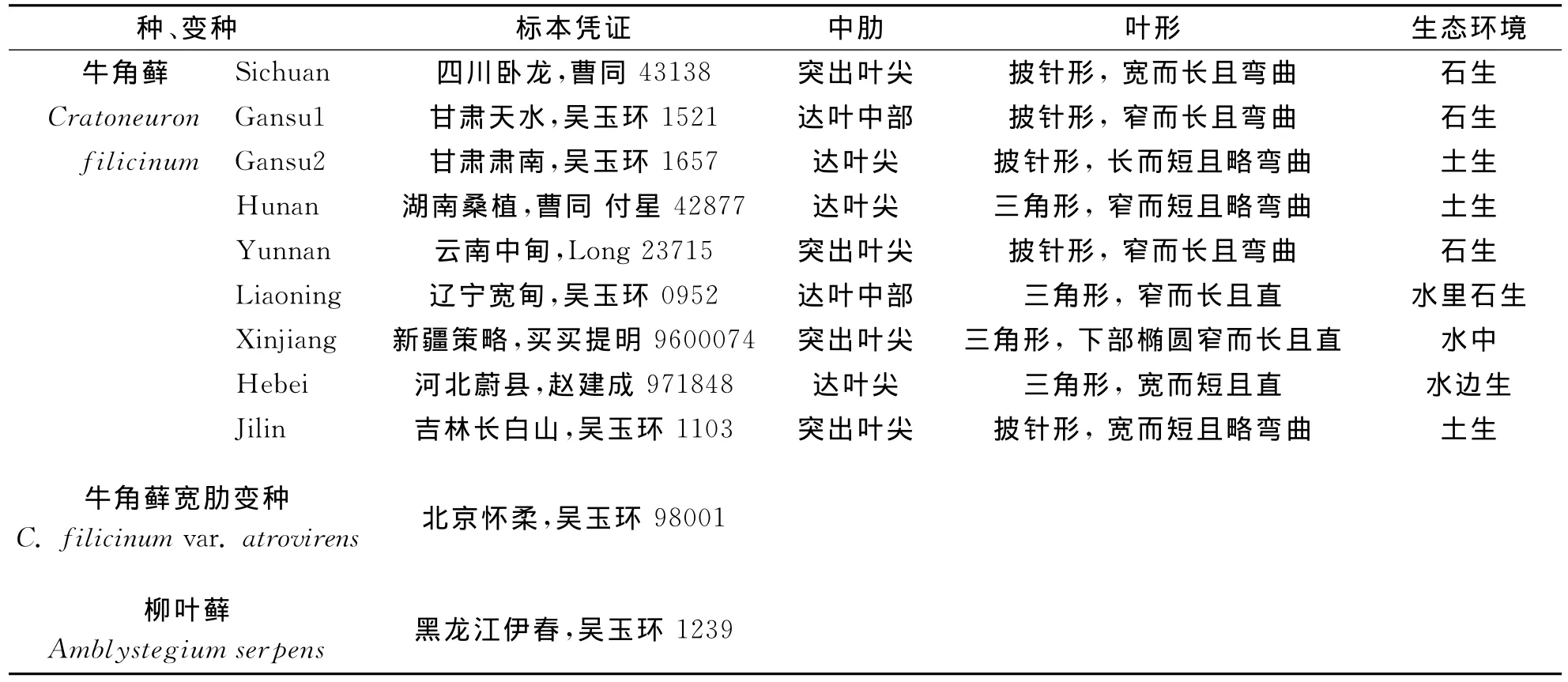
表1 试验用标本及其形态特征与采集地生态环境Tab.1 Specimens and their morphological characteristics and enviroments
1.2 方 法
1.2.1 基因组DNA提取(参考张道远等[12],稍作改动)
苔藓植物DNA提取过程中经常受到多酚和蛋白等物质的影响,难度较大.该研究中DNA的提取采用改良CTAB法,在研磨后加入含有PVP,Vc等抗氧化剂和稳定剂的缓冲液,能很好地抑制多酚氧化酶和细胞色素氧化酶的活性;抽提方法上,两次使用氯仿抽提,能有效地去除蛋白质等的影响.
将试验材料分别用70%酒精和蒸馏水清洗后放入研钵中,加入液氮迅速、充分研磨,待成极细粉末状时迅速转移至1.5mL离心管中,加入800μL冰浴预冷的DNA研磨缓冲液,充分混匀后2 000r/min 4℃离心10min,弃上清,在沉淀中加入600μL经65℃预热的裂解缓冲液,缓慢混匀,65℃水浴60min(中间不时轻轻搅动)后加入等体积氯仿,摇匀后6 000r/min 4℃离心10min.然后取上清液于1.5mL离心管中,再加入等体积氯仿,摇匀后6 000r/min 4℃离心10min.取上清液,加入1/5体积10mol/L NH4Ac和等体积预冷异丙醇,混匀后-20℃静止1h以上或过夜.12 000r/min 4℃离心10min沉淀DNA,再用70%乙醇洗涤沉淀数次,65℃干燥数分钟,将沉淀溶于30μL的TE中,-20℃保存.
DNA研磨缓冲液配方:0.1mol/L Tris-HCL,pH为8.0;5mmol/L Na2EDTA,pH为8.0;20mg/mL PVP;25mg/mLβ-巯基乙醇;1mg/mL Vc.
裂解缓冲液配方:20mg/mL CTAB;0.1mol/L Tris-HCl,pH为8.0;20mmol/L Na2EDTA,pH为8.0;1.4mol/L NaCl;20mg/mL PVP;25mg/mLβ-巯基乙醇;1mg/mL Vc.
1.2.2 RAPD扩增
反应体系主要成分为:10×PCR buffer 2.5μL(10mmol/L Tris-HCl,pH为8.3;50mmol/L KCl),模板DNA 100ng,dNTP 0.1mmol/L,Taq polymerase 0.75U,引物0.20μmol/L,Mg2+浓度2.00 mmol/L.反应体系总体积为25μL.RAPD反应程序为:94℃预变性3min;变性94℃1min,退火36.5℃1min延伸72℃1min35个循环;延伸72℃10min.冷却至4℃保存备用.
1.3 数据处理
PCR扩增反应重复1次,选取可重复的DNA带(包括强带和弱带)记录.每个样品的扩增带按“有”或“无”记录.“有”赋值为1,“无”赋值为0.根据Nei等[13]的方法计算样品间的简单遗传相似系数I,计算公式为I=2Nij/(Ni+Nj),其中,(Ni+Nj)为两样品的总扩增带数,Nij为两样品共有的扩增带数.利用MVSP程序,按非加权算术平均数聚类方法(UPGMA)计算种间遗传距离,建立系统聚类分析树状图.
2 结果与分析
2.1 遗传多态性
对60个引物进行筛选,得到30个多态性引物,从中挑选出15个条带清晰的引物进行统计分析(表2).这15个引物的G+C占64.4%,共扩增得到134条清晰稳定的扩增带,其中123条为多态性条带,多态条带比率(PPB)为91.79%,平均每个引物扩增出8.9条多态性带.片段大小在230~2 800bp之间,每个引物扩增6至14条不等,表明这些物种RAPD变异性较大,多态性高(图1).
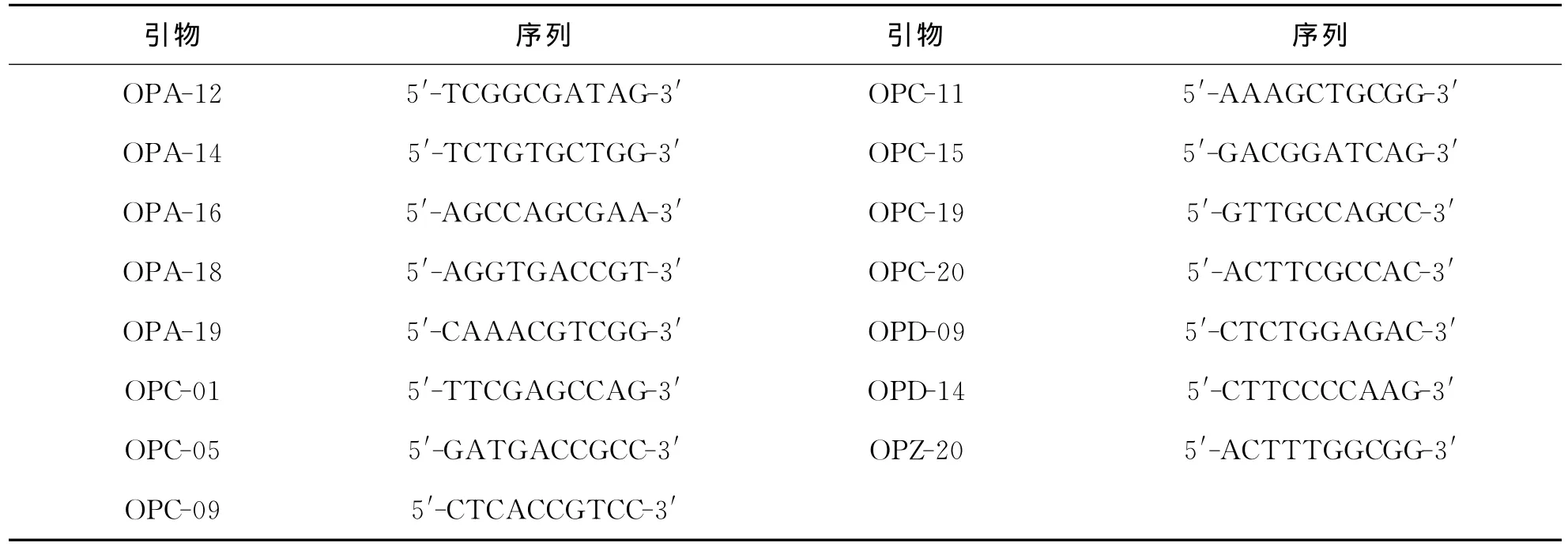
表2 RAPD分析用的15个随机引物序列Tab.2 Sequences of 15primers used by analysis of RAPD
2.2 遗传相似性
基于RAPD数据求得材料的遗传相似系数(GS),结果如表3所示.11份样品种内或种间两两之间的GS值在0.315~0.870之间,平均为0.714.供试材料的种内遗传相似系数较高,种间遗传相似系数较低:9份牛角藓样品两两之间的GS在0.703~0.870之间;而牛角藓宽肋变种与9份牛角藓样品的GS为0.617~0.683;柳叶藓与9份牛角藓样品的GS最低,在0.315~0.419之间.
2.3 牛角藓的聚类分析
供试材料的聚类分析结果见图2.从图中可以看出,11份样品可划分为3个类群:第一类群为柳叶藓;第二类群为牛角藓宽肋变种;第三类群为9份牛角藓样品.在不同的引物中,9份牛角藓样品都存在特有带,说明它们在基因组DNA的组成上具有共同序列.而这些带在牛角藓宽肋变种和柳叶藓中并不存在,说明这些条带是种的特征带,由此表明9份样品属同一种.此外,UPGMA聚类图显示9份牛角藓标本中产于北温带附近的6个聚在一起,而产于高纬度的2个牛角藓样品聚在一起.

图1 随机引物OPC-11扩增产生的RAPD带型Fig.1 RAPD patterns generated with random primers OPC-11

表3 基于RAPD数据计算的11份标本间的遗传相似系数Tab.3 Genetic similarity of 11specimens based on RAPD analysis
3 讨 论
该研究在分析每一个引物对试验样品扩增结果的基础上,将大量的DNA条带显示结果数字化,编入二进制输入计算机,得出每一样品的遗传相似性系数,以种内个体间相似系数的平均值为基础,得出种间聚类图.结果表明牛角藓遗传多样性较高,且表现为地域较近的样品首先聚合到一起,说明影响牛角藓遗传多样性的因素与所处地域的远近有关.牛角藓多生于水源处,基因流的转移有可能随着水流的变化而变化.从聚类图可以看出牛角藓与其变种的相似性较大而与柳叶藓较小,这是因为牛角藓与牛角藓宽肋变种为同属,亲缘关系较近,而与柳叶藓关系较远.
牛角藓为世界广布种,生于各种环境中,植物体大小和形态变化较大,尤其是叶片和细胞特征变化较大.对不同生态条件下采集的牛角藓标本进行形态特征观察,其叶片三角形或披针形,中肋或达于叶尖、或终于叶尖前部、或突出叶尖,叶片中部细胞从六边形到长菱形均有,表现出明显依生态环境尤其是固着基质和水湿条件不同而相异的特点.这些多变的生态型给系统分类带来许多争议.有研究[14]表明,苔鲜植物普遍存在着可塑性,其可塑性甚至比大多数有花植物还要强.这是因为在苔鲜植物中,选择作用有利于可塑性的产生,苔鲜植物有限的异体受精和基因交流能力限制了以产生新基因型的办法来提高对环境的适应能力[15].因此不应把形态特征上有明显变异的牛角藓简单地成立为新种,这些变异可能是环境饰变的结果.该研究中,虽然不同生长环境条件下的牛角藓植物体及其形态特征变化很大,但依据RAPD分析结果,9份牛角藓样品聚在一起,明显与外类群牛角藓宽肋变种和柳叶藓分离,充分说明其为同一物种.该结果为经典形态分类学的观点提供了新的分子证据.
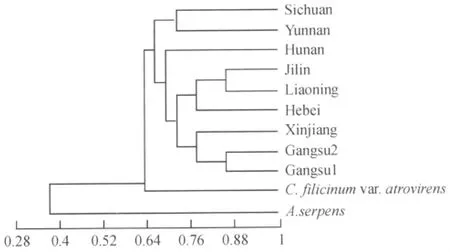
图2 基于RAPD数据用UPGMA法构建的树系图Fig.2 Dendrogram using UPGMA based on RAPD data
[1]Ochyra R.Animadversions on the moss genusCratoneuron(Sull.)Spruce[J].Journal of Hattori Botanical Laboratory,1989,67:203-242.
[2]Wu Yuhuan,Gao Chien,Li Wei.Amblystegiaceae[M]//Hu Renliang,Wang Youfang,Crosby M R.Moss Flora of China:Vol 7.Beijing:Science Press,2009:32-35.
[3]吴玉环,高谦,曹同.柳叶藓科[M]//胡人亮,王幼芳.中国苔藓志:卷7.北京:科学出版社,2005:6-12.
[4]Redfearn P L,Tan B C,He S.A newly updated and annotated checklist of Chinese mosses[J].Journal of Hattori Botanical Laboratory,1996,79:163-357.
[5]白学良,赵尊田.内蒙古苔藓志[M].呼和浩特:内蒙古大学出版社.1997:384-419.
[6]Wijk R,Margardant W D,Florschutz P A.Index Muscorum:Volume 1[M].Utrecht:International bureau for plant taxonomy and nomenclature of the international association for plant taxonmy,1959:1-548.
[7]吴玉环,罗昊,李微,等.牛角藓(Cratoneuronfilicinum(Hedw.)Spruce)叶片形态特征的生态变异及其分类学意义[J].生态学杂志,2007,26(6):882-885.
[8]刘丽,朱永青,王幼芳.鼠尾藓不同居群间形态及RAPD分析[J].云南植物研究,2006,28(6):570-574.
[9]Stenøien H K,Såstad S M.Genetic structure in three haploid peat mosses(Sphagnum)[J].Heredity,1999,82:391-400.
[10]Skotnicki M L,Selkirk P M,Broady P,etal.Dispersal of the mossCampylopuspyriformison geothermal ground near the summits of Mount Erebus and Mount Melbourne,Victoria Land,Antarctica[J].Antarctic Science,2001,13(3):280-285.
[11]Zhu Yongqing,Liu Li,Wang Youfang,etal.Genetic diversity and population structure ofBrachytheciumrivulareSchimp.(Brachytheciaceae)from Foping Nature Reserve,Shaanxi,China,detected by RAPD markers[J].Journal of Bryology,2007,29(2):104-110.
[12]张道远,张元明,曹同.耐旱苔藓植物DNA提取及优化RAPD、ISSR反应体系的建立[J].中国沙漠,2006,26(5):826-831.
[13]Nei M,Li W H.Mathematical model for studying genetic variation in terms of restriction endonucleases[J].Proc Natl Acad Sci USA,1979,76:5269-5273.
[14]Longton R E.Genecological differentiation in bryophytes[J].Journal of Hattori Botanical Laboratory,1974,38:48-65.
[15]崔明昆.苔藓植物的生态变异及其分类学中的生态学准则[J].云南师范大学学报:自然科学版,2001,21(4):61-64.
Study on the Classification of
Cratoneuronfilicinum(Hedw.)Spruce by RAPD
LI Peng-bo1,WU Yu-huan1,2,LUO Hao2
(1.College of Life and Envionmental Sciences,Hangzhou Normal University,Hangzhou 310036,China;2.Institute of Applied Ecology,Chinese Academy of Sciences,Shenyang 110016,China)
Choosing nineCratoneuronfilicinumspecimens collected from Sichuan,Gansu,Hunan,Yunnan,Liaoning,Hebei,Jilin and Xinjiang as the materials,the paper researched on the taxonomic revision of differentC.filicinumby RAPD,obtained 134distinct bands from 15primers,of which 91.79%were polymorphic,and the genetic similarity coefficient(GS)among nineC.filicinumspecimens varied from 0.703to 0.870.The results showed that the individuals of nine specimens are the same specie though they are very different in morphological characteristics.
Cratoneuronfilicinum;RAPD;taxonomic revision;environment modification
Q949
A
1674-232X(2011)03-0237-05
10.3969/j.issn.1674-232X.2011.03.010
2010-12-28
国家自然科学基金项目(30770161,30970188);杭州师范大学科研启动基金项目(YS05203130).
李鹏勃(1986—),男,陕西周至人,植物学专业硕士研究生,主要从事植物系统进化与生态学研究.
*通信作者:吴玉环(1972—),女,浙江苍南人,教授,博士,主要从事植物系统进化与生态学研究.E-mail:yuhuanwu@yahoo.com.cn
