虾夷扇贝生殖腺多肽的制备及分离
2011-09-26金文刚,吴海涛,朱蓓薇,,刘冰,许景光
金 文 刚, 吴 海 涛, 朱 蓓 薇,, 刘 冰, 许 景 光
( 1.西北农林科技大学 食品科学与工程学院, 陕西 杨凌 712100;2.大连工业大学 食品学院, 辽宁 大连 116034 )
0 引 言
扇贝柱产品在生产和加工过程中,会产生大量的低值副产物,占扇贝湿重的70%左右,包括贝壳(52%)、裙边(9%)、中肠腺(5%)、性腺(卵巢和精巢,3%~9%)[1]。研究表明,扇贝加工副产物中含有大量蛋白质[2]、多糖[3]及脂类[4]等成分,对扇贝加工副产物进行开发利用,不仅能减少生物资源的浪费,还可避免由此引起的环境污染。
利用生物酶制备生物活性肽,是当前海产品副产物蛋白质回收利用的有效途径之一,具有安全性高、条件温和、易于控制、对设备要求低等优点。有关利用扇贝加工副产物中蛋白资源的研究,主要是以扇贝裙边或内脏团为原料通过外源酶水解,制备调味料[5]或活性肽[6]。扇贝生殖腺是扇贝内脏团的重要组成部分,也是鲜食扇贝的可食用部分,具有较高的安全性,国外对扇贝生殖腺中的酶[7]、脂肪酸[8]和类胡萝卜素[9]等进行了相关研究,然而,采用外源酶对扇贝生殖腺进行酶解,制备多肽的研究还鲜见报道。为此,本研究分别以雌、雄虾夷扇贝生殖腺为原料,结合变性预处理,选用中性蛋白酶制备虾夷扇贝生殖腺多肽,比较雌性和雄性虾夷扇贝生殖腺的酶解效果及水解液的组成,并对雌性虾夷扇贝生殖腺水解液中多肽进行了初步分离,考察其还原能力及分子质量分布情况,为虾夷扇贝生殖腺的高值化利用提供理论依据。
1 材料与方法
1.1 材料与试剂
试验材料:鲜活虾夷扇贝,购于大连长兴水产品批发市场,低温运至实验室后,立即从雌性和雄性扇贝内脏团中摘下生殖腺,其中雌性生殖腺呈橘红色,雄性生殖腺呈乳白色,将其冲洗后分装,-20 ℃冷冻保藏备用。
主要试剂:中性蛋白酶,南宁庞博生物制药有限公司;Sephadex G-25葡聚糖凝胶,Pharmacia公司;Cytochrome C(12.5 ku),Aprotinin(6.512 ku),上海生工生物工程技术服务有限公司;18肽(2.342 ku),西安联美生物科技有限公司;Vitamin B12(1.355 ku),N-Hippuryl-His-Leu(0.43 ku),Sigma公司;其余试剂为国产分析纯。
1.2 仪器与设备
pHS-3C型精密pH计,上海雷磁仪器厂;UV2100型紫外可见分光光度计,上海尤尼柯仪器有限公司;BSE-100型自动部分收集器,上海沪西分析仪器厂;P230p型高效液相色谱,大连依利特分析仪器有限公司;SuperdexTM Peptide 10/300GL凝胶柱,GE Healthcare公司。
1.3 方 法
1.3.1 虾夷扇贝生殖腺主要化学组成分析
水分的质量分数采用常压干燥法测定;粗蛋白的质量分数采用微量凯氏定氮法测定;粗脂肪的质量分数采用索氏抽提法测定;总糖的质量分数采用苯酚硫酸法,以葡萄糖为标准品,所得标准曲线为y=0.012 6x-0.018 2(R2=0.992 8);还原糖的质量分数采用DNS法,以葡萄糖为标准品,所得标准曲线为y=0.001x+0.001(R2=0.999 4)。
1.3.2 酶解方案
1.3.2.1 变性预处理
雌、雄虾夷扇贝生殖腺分别经匀浆后,加水至底物质量分数达到4%,沸水浴10 min进行变性预处理(变性处理组),并在加热过程中保鲜膜封口,防止水分流失,试验中以未经变性预处理的样品为对照(未变性处理组)。
1.3.2.2 酶解条件
选用中性蛋白酶(试验前测定的酶活为64 812 U/g),以4%蛋白质量分数的生殖腺匀浆为底物,酶加量为3 000 U/g蛋白,pH为7.0,温度为50 ℃,酶解时间为3 h。
1.3.2.3 酶解工艺
准确称取雌、雄虾夷扇贝生殖腺匀浆→分别补水至100 mL(底物质量分数达到4%)→分组(变性处理和未变性处理)→调pH至7.0→加酶→50 ℃恒温酶解3 h→灭酶(100 ℃,10 min)→离心(4 000 r/min,15 min)→上清液分装后冷冻保藏。
1.3.3 水解度测定
采用pH-stat法,以滴定所消耗的标准NaOH溶液体积计算水解度。
1.3.4 水解液中可溶蛋白、TCA可溶性寡肽质量浓度和肽得率的测定
将经变性处理和未变性处理的雌、雄虾夷扇贝生殖腺水解液稀释一定倍数后,分别测定水解液中可溶蛋白、TCA可溶性寡肽质量浓度以及肽得率。可溶蛋白质量浓度采用考马斯亮蓝 G-250法,以牛血清蛋白为标准,所得标准曲线为y=0.009 9x(R2=0.991 8);TCA可溶性寡肽的质量浓度以及肽得率采用三氯乙酸(TCA)沉淀结合Folin-酚的方法,分别取水解液2 mL于5 mL离心管中,加入20%的TCA溶液2 mL,静置20 min,4 000 r/min离心15 min,取1 mL稀释一定倍数的上清液,采用Folin-酚法测定样品中可溶性寡肽质量浓度。以牛血清蛋白为标准品,绘制的标准曲线为y=0.003 5x+0.001 28(R2=0.997 7)。根据式(1)计算肽得率:
肽得率=m1/m0
(1)
式中,m1为水解液中TCA可溶性寡肽质量,mg;m0为水解液中总蛋白质量,即水解液中底物蛋白量。
1.3.5 雌性虾夷扇贝生殖腺水解液Sephadex G-25 凝胶层析分离及还原能力测定
Sephadex G-25凝胶层析柱(1.6 cm×50 cm),去离子水平衡12 h,雌生殖腺水解液上样量为0.5 mL,以去离子水为洗脱液,采用部分收集器收集,体积流量为每20 min 5 mL,每次收集40管,分别测定OD280及还原能力。
还原能力测定参照文献[10]并作修正。取样品溶液1 mL,依次加入1 mL的磷酸盐缓冲液(0.2 mol/L,pH 6.6),2.0 mL 1%铁氰化钾[K3Fe(CN)6]溶液,混匀,在50 ℃下保温20 min。再加入10%的三氯乙酸溶液1 mL,振荡混匀后,4 000 r/min离心10 min。取上清液2 mL,加入2.5 mL蒸馏水和0.3 mL 0.1%的三氯化铁(FeCl3)溶液,静置10 min,测定OD700,空白组以等体积去离子水代替样品溶液。
1.3.6 变性处理雌生殖腺水解液的分子质量分布
变性处理雌生殖腺水解液稀释一定倍数,过0.45 μm微孔膜后,分子质量分布采用依利特P230高效液相色谱结合Superdex Peptide 10/300 GL(10 mm×300 mm)凝胶色谱柱测定。进样量:50 μL;检测波长:220 nm;流动相:0.05 mol/L磷酸盐缓冲液(含0.15 mol/L NaCl,pH 7.0);洗脱流量:0.5 mL/min。分子质量标准品分别选用Cytochrome C(12. 5 ku),Aprotinin(6.512 ku),18肽(2.342 ku),Vitamin B12(1.355 ku)、N-Hippuryl-His-Leu(0.43 ku)和gly-gly (0.132 ku)。对凝胶过滤色谱而言,分子质量的对数(y)与保留时间(x)呈线性关系,经过计算所得分子质量标准曲线为y=-0.083 4x+5.755 4(R2=0.994 4)。
2 结果与讨论
2.1 虾夷扇贝生殖腺的主要化学组成
经测定,雌、雄虾夷扇贝生殖腺原料的水分质量分数分别为(82.63±0.05)%和(79.70±0.02)%,生殖腺中粗蛋白、粗脂肪、总糖、还原糖的质量分数见表1。由表1可见,虾夷扇贝生殖腺的主要成分为蛋白质,与雌生殖腺相比,雄生殖腺的蛋白质量分数相对较大,而雌生殖腺脂肪的质量分数大于雄生殖腺。Palacios等[11]指出雌生殖腺在配子形成期易于富集脂肪,本研究采用的虾夷扇贝生殖腺在4月份生殖腺指数较高,这可能是雌、雄生殖腺脂肪质量分数差异较大的原因;雌、雄生殖腺中总糖、还原糖的质量分数无显著差异,Ruiz等[12]发现不同季节的生殖腺,蛋白、脂肪和碳水化合物含量存在差异,他们用Bradford法测定的欧洲大扇贝生殖腺中蛋白质量分数在61.2~187.3 mg/g,脂肪和碳水化合物质量分数在6.7~20.6 mg/g和14.1~33.2 mg/g,与本研究测定的虾夷扇贝生殖腺的化学组成结果基本一致。
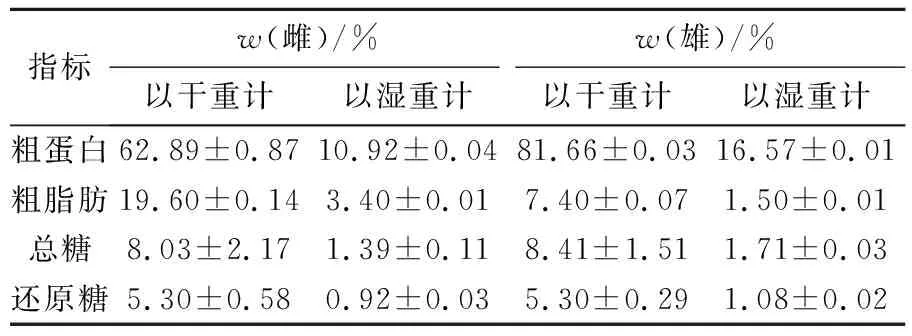
表1 虾夷扇贝生殖腺化学成分的质量分数(4月)
2.2 虾夷扇贝生殖腺酶解过程中的水解度变化
对虾夷扇贝生殖腺进行变性预处理,以未变性组为对照,雌、雄虾夷扇贝生殖腺经中性蛋白酶酶解,水解度随水解时间变化曲线见图1。由图1可见,随着水解时间的延长,雌、雄生殖腺变性处理组和未变性处理组的水解度在30 min前显著上升,之后水解度逐渐趋于平缓,至3 h达到平 台期,雌生殖腺未变性和变性处理组的水解度分别达到25.08%和34.15%;雄生殖腺未变性和变性处理组的水解度分别达到18.37%和28.21%。可见,中性蛋白酶水解雌性生殖腺效果优于雄性生殖腺,并且底物经变性处理后,雌、雄生殖腺的水解度分别提高了36.16%和53.57%。究其原因可能是由于虾夷扇贝生殖腺蛋白质经变性处理后,蛋白质空间结构,即二、三、四级结构被破坏,中性蛋白酶与蛋白质分子内部的酶切位点更容易发生作用,从而促进了酶解,这与于志鹏等[13]发现蛋清酶解前热变性处理得到水解度增大的结果类似。

图1 虾夷扇贝生殖腺酶解过程中的水解度曲线
2.3 虾夷扇贝生殖腺水解液中可溶蛋白、TCA可溶性寡肽质量浓度及肽得率
经测定,雌、雄生殖腺变性处理组和未变性处理组的水解液中,可溶蛋白、TCA可溶性寡肽质量浓度及肽得率结果见表2。由表2可知,与雄生殖腺相比,雌生殖腺水解液中可溶性蛋白质量浓度较低,并且变性处理组的可溶蛋白的质量浓度也低于未变性处理组,而雄生殖腺变性处理和未变性组水解液中,可溶性蛋白的质量浓度差异不大;雌、雄生殖腺变性处理组的水解液,TCA可溶性寡肽质量浓度以及肽得率均高于未变性处理组,其中雌、雄生殖腺变性处理组水解液中,TCA可溶性寡肽质量浓度分别为未变性处理组的2.32和1.05倍,而雌、雄生殖腺变性处理组水解液中,肽得率分别为未变性处理组的1.95和1.27倍。以上结果表明,以雌性虾夷扇贝生殖腺为底物,利用中性蛋白酶结合变性处理工序,能获得较高多肽得率的水解液,酶解效果较好。
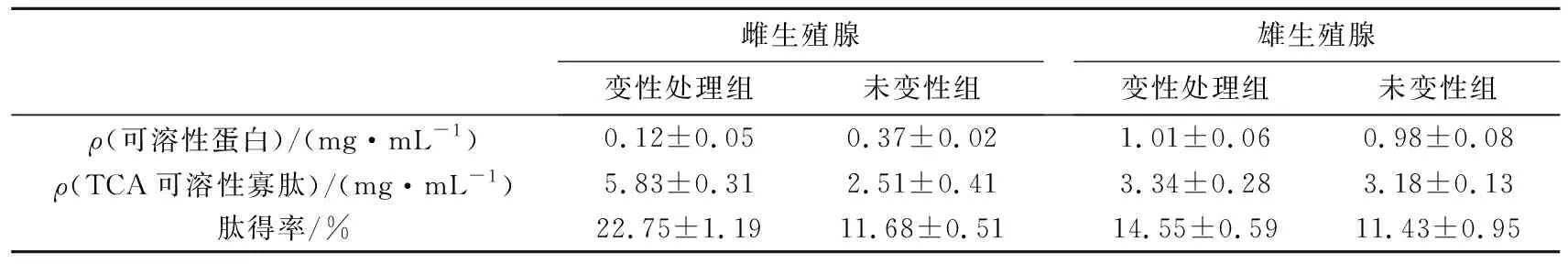
表2 虾夷扇贝生殖腺水解液中可溶性蛋白、TCA可溶性寡肽质量浓度及肽得率
2.4 雌性虾夷扇贝生殖腺水解液的Sephadex G-25凝胶层析分离及还原能力
以中性蛋白酶为外源酶,对雌性虾夷扇贝生殖腺进行酶解,可获得较高肽得率的水解液。进一步将雌生殖腺变性处理和未变性处理组的水解液,经Sephadex G-25凝胶层析分离,并监测其还原能力,结果见图2。 由图2可见,未变性处理的雌生殖腺水解液经Sephadex G-25凝胶层析分离后,获得4个主要蛋白峰,洗脱液在280 nm的吸光值分别在第10、16、24及30管出现峰值,其中第1、2、4峰都具有一定的还原能力,表明组分可能有一定的抗氧化活性[10];经变性处理的雌生殖腺水解液经Sephadex G-25凝胶分离后,仅获得2个主要蛋白组分,洗脱液在280 nm的吸光值分别在第9及16管出现峰值,二者均有一定的还原能力,表明变性处理后雌生殖腺水解液比未变性处理的水解液的分子质量分布更为均一。
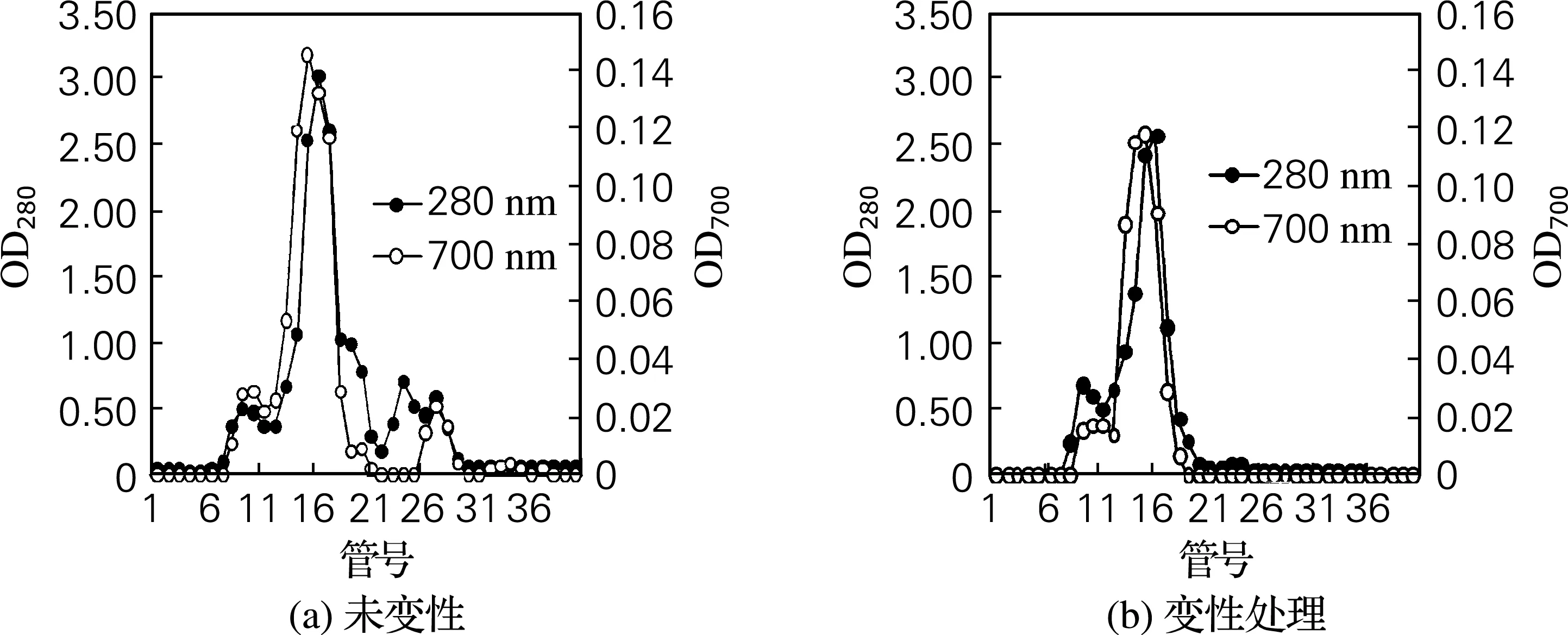
图2 雌性虾夷扇贝生殖腺水解液的Sephadex G-25凝胶层析分离及还原能力
2.5 经变性处理后雌性虾夷扇贝生殖腺水解液的分子质量分布
对雌性生殖腺变性处理组水解液的分子质量分布进行考察,结果见图3。根据保留时间并结合峰面积,对照标准品的分子质量和保留时间,发现雌生殖腺变性处理组水解液的分子质量分布范围主要集中于0.25~5 ku,占水解液的74.78%,符合已报道的活性多肽的分子质量范围。此外,研究还发现,雄性虾夷扇贝生殖腺经中性蛋白酶水解后,保水效果较好,在今后的研究中将对其理化性质进行研究。
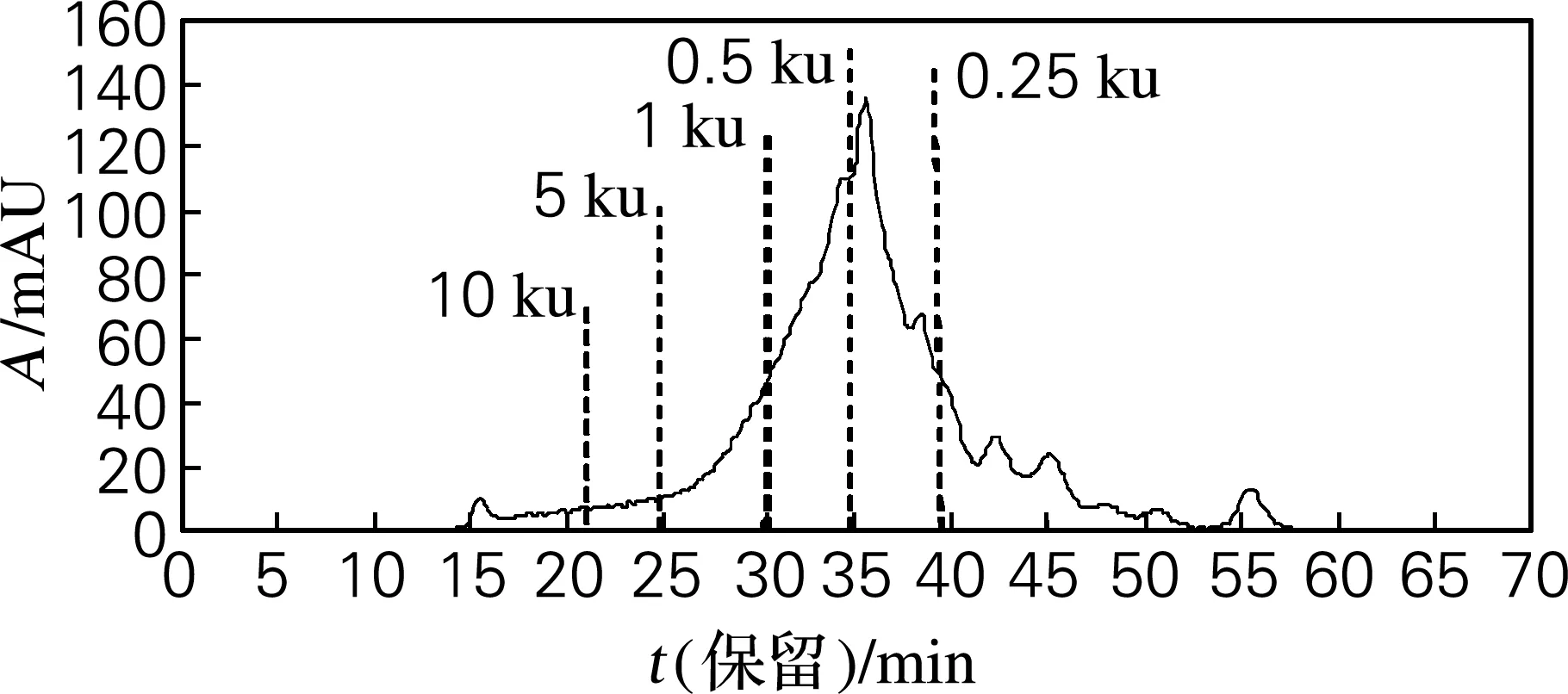
图3 雌性虾夷扇贝生殖腺变性处理组水解液中肽的分子质量分布
3 结 论
虾夷扇贝生殖腺中粗蛋白的质量分数分别达到干重的(62.89±0.87)%(雌性)和(81.66±0.03)%(雄性),是制备活性肽的良好来源。利用中性蛋白酶对虾夷扇贝生殖腺进行酶解,雌性生殖腺水解效果优于雄性生殖腺。虾夷扇贝生殖腺经煮沸变性处理后,可显著提高水解液的水解度和肽得率。结合变性预处理工序,采用中性蛋白酶对雌性虾夷扇贝生殖腺进行酶解,可获得低分子质量的多肽,分子质量分布范围主要集中于0.25~5 ku,占水解液的74.78%。Sephadex G-25凝胶层析可以对雌性虾夷扇贝生殖腺水解液进行分离,可获得具有还原能力的多肽组分。
[1] OYAMADA C, KANENIWA, M, EBITANI K, et al. Mycosporine-like amino acid extracted from scallop (Patinopectenyessoensis) ovaries: UV protection and growth stimulation activities on human cells[J]. Marine Biotechnology, 2008, 10:141-150.
[2] 郑丽,汪秋宽. 扇贝加工废弃物蛋白酶解及其酶解产物分子量分布的研究[J]. 大连水产学院学报, 2006, 21(2):149-152.
[3] ZHU Beiwei, ZHOU Dayong, YANG Jingfeng, et al. Structural analysis of a polysaccharide fromPatinopectenyessoensisviscera[J]. European Food Research and Technology, 2009, 229:971-974.
[4] ZHOU Dayong, ZHU Beiwei, TONG Lei, et al. Extraction of lipid scallop (Patinopectenyessoensis) viscera by enzyme-assisted solvent and supercritical carbon dioxide methods[J]. International Journal of Food Science and Technology, 2010, 45(9):1787-1793.
[5] 郑丽,汪秋宽. 扇贝加工废弃物海鲜调味料的加工利用[J]. 水产科学, 2005, 24(1):34-37.
[6] 胡文婷,孙谧,王跃军. 栉孔扇贝(Chlamysfarreri)中抗氧化肽的分离纯化及性质研究[J]. 海洋与湖沼, 2006, 37(1):14-19.
[7] TERENT’EV L L, TERENT’EVA N A, RASSKAZOV V A. Thymidine and thymidylate kinases from the scallopMizuhopectenyessoensisgonads[J]. Applied Biochemistry and Microbiology, 2008, 44(5):515-522.
[8] SOUDANT P, MARTY Y, MOAL J, et al. Effect of food fatty acid and sterol quality onPectenmaximusgonad composition and reproduction process[J]. Aquaculture, 1996, 143:361-378.
[9] SUHNEL S, LAGREZE F, FERREIRA J F, et al. Carotenoid extraction from the gonad of the scallopNodipectennodosus(Linnaeus, 1758) (Bivalvia: Pectinidae)[J]. Brazilian Journal of Biology, 2009, 69(1):209-215.
[10] ZHU Lijuan, CHEN Jie, TANG Xueyan, et al. Reducing, radical scavenging, and chelation properties ofinvitrodigests of alcalase-treated zein hydrolysate[J]. Journal of Agricultural and Food Chemistry, 2008, 56(8):2714-2721.
[11] PALACIOS E, RACOTTA I S, KRAFFE E, et al. Lipid composition of the giant lion’s-paw scallop(Nodipectensubnodosus) in relation to gametogenesis I. Fatty acids[J]. Aquaculture, 2005, 250:270-282.
[12] RUIZ-VERDUGO C A, RACOTTA I S, IBARRA A M. Comparative biochemical composition in gonad and adductor muscle of triploid and diploid catarina scallop (ArgopectenventricosusSowerby II, 1842)[J]. Journal of Experimental Marine Biology and Ecology, 2001, 259:155-170.
[13] 于志鹏,林松毅,刘静波,等. 优化蛋清粉蛋白热变性条件的研究[J]. 食品科学, 2007, 28(10):229-232.
