大贵寺国家森林公园野生青檀群落结构及物种多样性
2011-06-12汪殿蓓李建华田春元刘仁阳
汪殿蓓,李建华,田春元,刘仁阳
(孝感学院生命科学技术学院,湖北 孝感 432000)
0 引 言
青檀(Pteroceltis tatarinowii Maxim.)为榆科青檀属落叶乔木,是中国特有的单种属植物及特有的纤维树种,树皮纤维是制造宣纸的主要原料,木材坚硬致密,是制作农具、车轴、家具、建筑用的上等木料[1].近年来,对青檀的研究报道集中于其栽培林的培育措施及机理:探讨了青檀种子的休眠机理[2]、种子的活力[3]、盐胁迫对青檀种子萌发及幼苗生长的影响[4]、一年生播种苗生长发育规律[5]、叶的解剖结构及其生态适应性特征[6]、立地条件[7-8],经营措施[9],成土母岩和条龄[10-11]对青檀人工林生产力(生物量)、檀皮产量及檀皮中矿质元素含量的影响等.
青檀分布于我国安徽、浙江、福建、湖北等19省,常生于山谷溪边石灰岩山地疏林中[1].不过,由于自然植被破坏及大量砍伐,青檀分布区在逐渐缩小,野生群落面积越来越小,迫切需要对青檀野生群落进行研究,阐明野生青檀群落结构及变化规律,为针对性保护措施的制定提供依据.目前对青檀天然群落结构及多样性的研究较少报道[12-14].
湖北大贵寺国家森林公园青檀群落是分布面积较大、保存较好的野生群落,本文以该野生青檀群落为研究对象,研究其物种组成、区系成分、物种多样性等内容,以促进青檀野生种质资源的保护和利用.
1 研究地概况及研究方法
1.1 研究地概况
大贵寺国家森林公园位于湖北省广水市北35 km(113°31′~114°07′ E,31°23′~32°05′ N),北面距河南信阳37 km.属低山丘陵地带,公园的最高点——大贵寺金顶海拔908 m.为北亚热带大陆性季风气候,年均气温13~14 ℃,年均降水量990 mm;平均无霜期221 d,全年日照数2 083 h.地带性植被为常绿落叶阔叶混交林[15].
1.2 研究方法
1.2.1 野外调查 采用典型取样法在青檀集中分布处设置样方进行群落调查,共设置10 m×10 m样方29个.胸径≥2 cm的树种划入乔木层,对样方中的乔木种进行每木检测,记录种类、胸径、高度,调查记录灌木的种类、数量、高度、冠幅.在10 m×10 m样方中设置5个1 m×1 m小样方,在小样方中调查草本植物种类、数量、平均高度和盖度.各个样方的生境条件如表1.
1.2.2 数据处理 按Raunkiaer植物生活型分类系统统计群落植物生活型谱,计算乔木层、灌木层、草本层各层次物种的重要值[16].按吴征镒种子植物属分布区类型划分方法分析属的分布类型及其区系组成[17-18].采用物种丰富度指数、Shannon-wiener (Shannon- Weaver)、Simpson指数和Pielous均匀度指数[16]计算群落物种多样性.
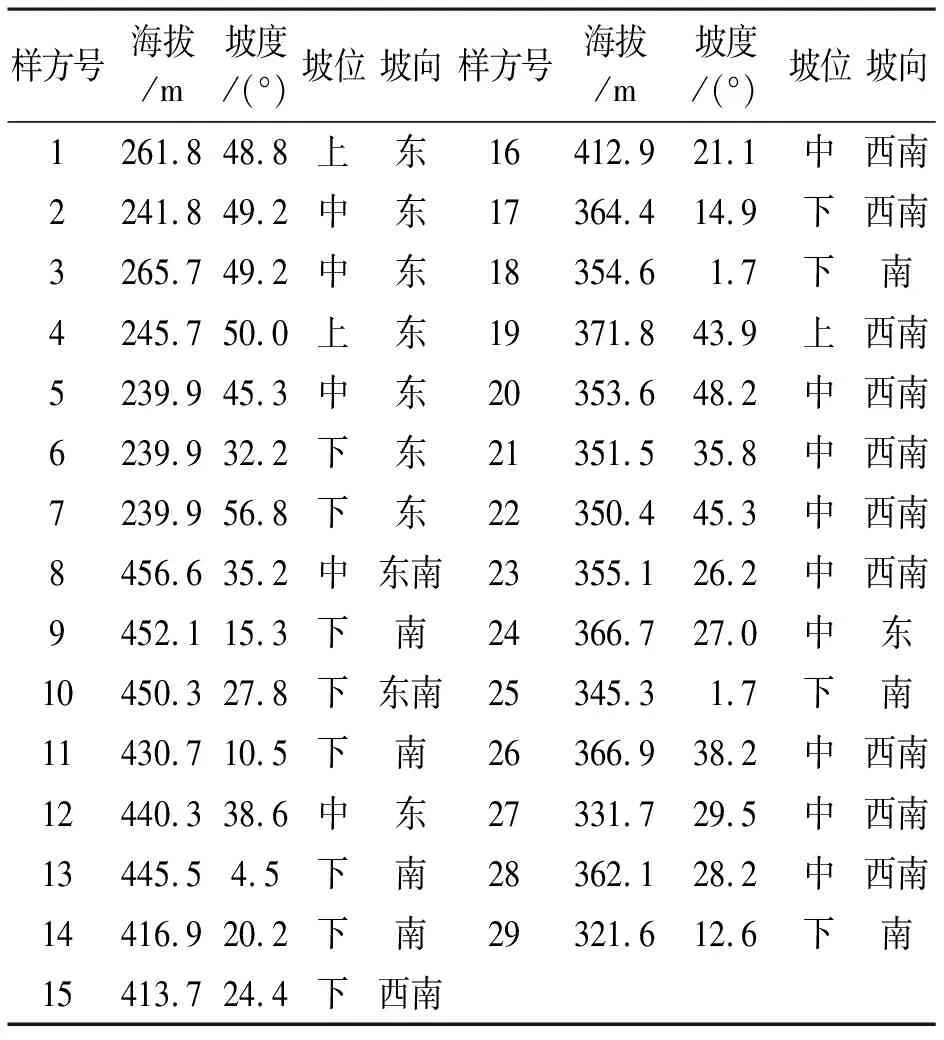
表1 各个样方的生境条件
2 结果与分析
2.1 群落生活型谱
植物生活型是群落结构和外貌的重要组成部分.研究植物生活型不仅可以了解区域的生物气候特征,还可为群落分类及结构特征的揭示等提供重要的依据.一个群落的组成种可以划分出几种生活型,但在一定气候条件下总是以特定的生活型谱为特征[19].表2为青檀群落植物生活型谱,其中高位芽植物84种,占总数的53.8%,常绿高位芽植物有18种,占11.5%,落叶高位芽植物66种,占42.3%;地上芽植物7种,占4.5%;地面芽植物36种,占23.1%;地下芽植物15种,占9.6%;一年生植物14种,占9.0%.该生活型谱组成反映出了地理环境对群落的影响,该青檀群落地处湖北北部,与河南省接壤,属北亚热带区域,因此群落中仍以高位芽占一定优势,但与中亚热带地区群落[20]、南热带地区植物群落[21]相比,高位芽比例减少,地面芽植物的比例增大,与冬季低温的影响有关.
表2青檀群落植物生活型谱
Table 2 Life form spectrum of P. tatarinowii community
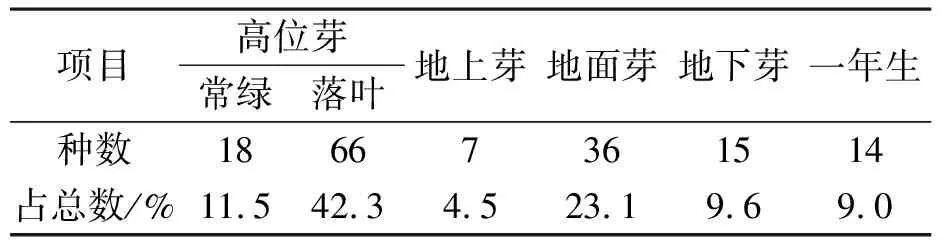
项目高位芽常绿落叶地上芽地面芽地下芽一年生种数18667361514占总数/%11.542.34.523.19.69.0
2.2 群落的种类组成
2 900 m2样地中有植物156种,分属60科108属,其中种子植物55科103属149种,蕨类植物5科5属7种,群落种类组成较丰富.种类较多的科为百合科(8属13种),蔷薇科(5属8种),豆科(7属7种),壳斗科(3属8种),榆科(4属8种),桑科(4属7种),樟科(3属7种)等.
乔木层共有植物49种,以青檀为优势种,重要值为155.9,其次是小叶朴(CeltisbungeanaBlume)和锐齿槲栎(Quercusalienavar.acuteserrataMaxim.ExWenz.),重要值分别为13.5和11.7,重要值有前10位的种还有朴树(CeltissinensisPers.)、杉木(Cunninghamialanceolata(Lamb.) Hook)、豹皮樟(Litseacoreanavar. sinensis(Allen) Yang et P. H. Huang)、油桐(Verniciafordii(Hemsl.) Airy Shaw)、山樱桃(PrunusserrulataLindl.)、臭椿(Ailanthusaltissima(Mill.) Swingle)、榆树(UlmuspumilaL.).灌木层植物84种,包括乔木的幼树和真正的灌木种类,以青檀幼树为优势种,重要值为58.8,其次是锐齿檞栎、建始槭(AcerhenryiPax)、山胡椒(Linderaglauca(Sieb. Et Zucc.) Bl.)等,重要值分别为16.1、15.9、14.8,除建始槭、南方六道木(Abeliadielsii(Graebn.) Rehd.)、野桐(Mallotusjaponicus(Thunb.) Muell. Arg. var.floccosus(Muell. Arg) S. M. Hwang)等为真正的灌木种外,该层次中很多为乔木幼树构成.草本层植物44种,重要值在前10位的物种分别是贯众(CyrtomiumfortuneiJ.)、紫萁(OsmundajaponicaThunb.)、黑足鳞毛蕨(Dryopterislabordei(Christ)C.Chr.)、沿阶草(OphiopogonbodinieriLévl.)、野青茅(Deyeuxiaarundinacea(Linn.) Beauv.)、荞麦叶大百合(Cardiocrinumcathayanum(Wilson) Stearn)、紫堇(CorydalisedulisMaxim.)、阔叶山麦冬(LiriopepalatyphyllaWang et Tang)、井栏边草(PterismultifidaPoir.)和深山堇菜(ViolaselkirkiiPursh.),重要值分别为44.9,29.6,28.2,25.1,21.4,17.3,13.8,12.6,8.9,8.0.
藤本植物18科22属27种,主要由毛茛科、葡萄科、茜草科、蔷薇科等科植物组成,如毛茛科铁线莲属的铁线莲(ClematisfloridaThunb.)、单叶铁线莲(ClematishenryOliv.),葡萄科爬山虎属的爬墙虎(ParthenocissustricuspidataPlanch.)、绿爬墙虎(ParthenocissuslaetivirensRehd.),茜草科茜草属的茜草(RubiacordifoliaLinn.),蔷薇科的喜阴悬钩子(RubusmesogaeusFocke),菝契科菝契属的菝契(SmilaxchinaL.)、牛尾菜(SmilaxripariaA.DC.),五加科常春藤属的常春藤(HederanepalensisK. Koch. var.sinensis(Tobl.) Rehd.),葫芦科绞股蓝属的绞股蓝(Gynostemmapentaphyllum(Thunb.) Makino),夹竹桃科络石属的络石(Trachelospermumjasminoides(Lindl.) Lem.)等.
2.3 种子植物属区系成分分析
根据吴征镒[17-18]的《中国种子植物属的分布区类型》,可以将青檀群落种子植物103个属划分为13个分布型6个变型(表3).
表3青檀群落种子植物属分布区类型及变型
Table 3 The areal-types and subtypes of genera of seed plants in P. tatarinowii community
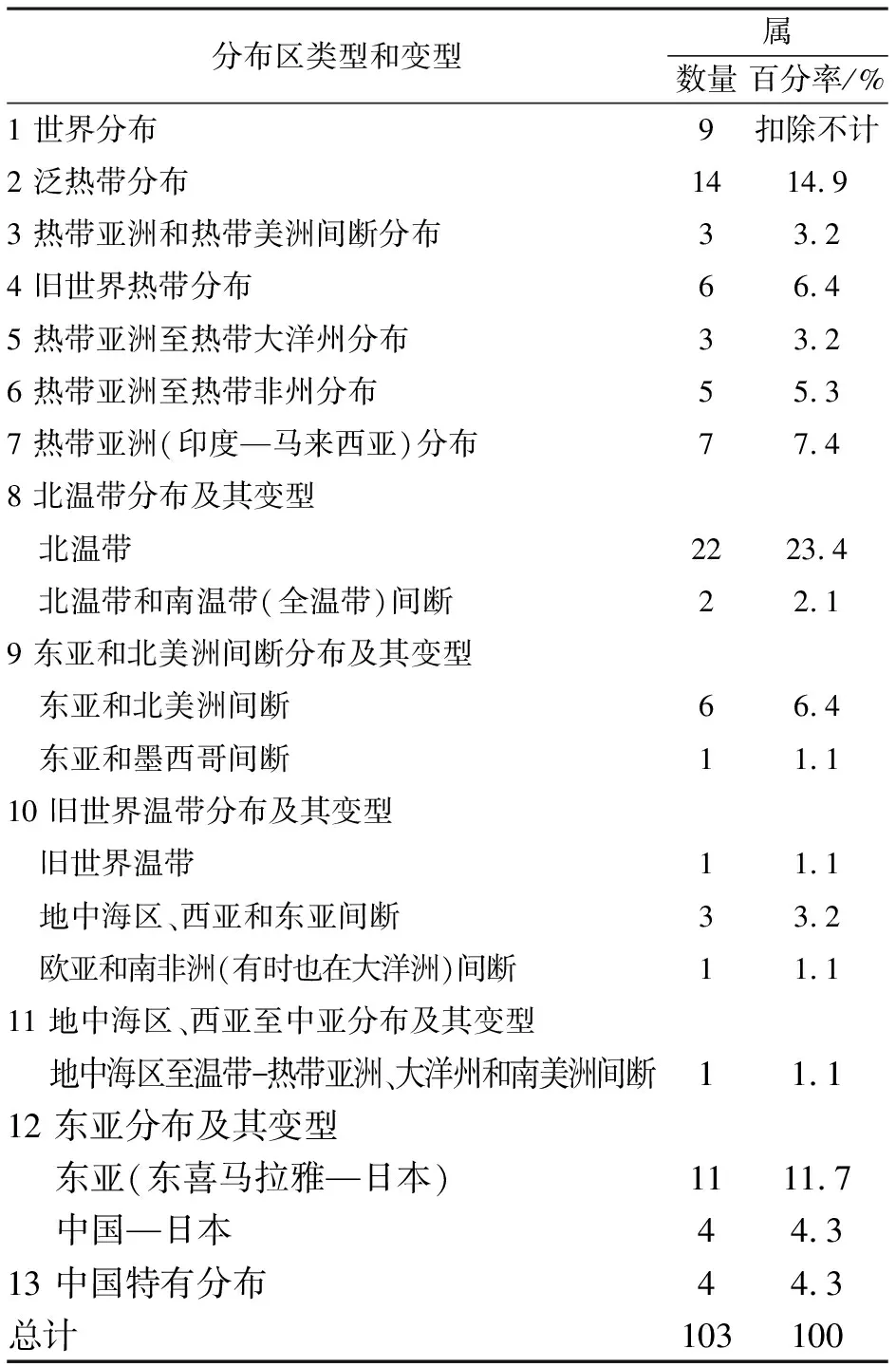
分布区类型和变型属数量百分率/% 1 世界分布9扣除不计2 泛热带分布1414.93 热带亚洲和热带美洲间断分布 33.24 旧世界热带分布 66.45 热带亚洲至热带大洋州分布33.26 热带亚洲至热带非州分布55.37 热带亚洲(印度—马来西亚)分布 77.48 北温带分布及其变型 北温带2223.4 北温带和南温带(全温带)间断22.19 东亚和北美洲间断分布及其变型 东亚和北美洲间断66.4 东亚和墨西哥间断 11.110 旧世界温带分布及其变型 旧世界温带11.1 地中海区、西亚和东亚间断 33.2 欧亚和南非洲(有时也在大洋洲)间断11.111 地中海区、西亚至中亚分布及其变型 地中海区至温带-热带亚洲、大洋州和南美洲间断 11.1 12 东亚分布及其变型 东亚(东喜马拉雅—日本)1111.7 中国—日本 44.3 13 中国特有分布 44.3 总计103100
2.3.1 世界分布 该分布区类型有9属,包括蓼属(PolygonumLinn.)、槐属(SophoraLinn.)、悬钩子属(RubusLinn.)、铁线莲属(ClematisLinn.)、堇菜属(ViolaLinn.)、千里光属(SenecioLinn.)等.由于世界分布属的生态适应幅度广,很难看出植物区系的地理特点,故在统计分析时被扣除.世界分布属大多是进化地位较高、对环境适应力较强、属内种数较多的属,在一定程度可反映出某一区域植物区系的进化程度、生态环境的特异性、人类活动的历史及其影响强度.如果人类活动时间较长,对生境破坏程度就越严重,本土植物就会减少,世界分布属所占的比例将增高.青檀群落世界分布属占中国种子植物世界分布属(104属)总数的8.7%,说明该群落存在着一定的人类活动干扰.
2.3.2 热带分布 是指分布中心或原始类型在南、北两半球热带地区的属,其绝大部分属起源于古南大陆,只有热带亚洲分布类型中有一部分起源于古老大陆北部,包括泛热带分布、热带亚洲和热带美洲间断分布、热带亚洲(印度-马来西亚)等类型[17].大贵寺森林公园青檀群落有热带分布属38 属,占总属数的40.4%.
其中泛热带分布14属,在热带分布属中数量最多,如榕属(FicusLinn.)、柿属(DiospyrosLinn.)、黄檀属(DalbergiaLinn.)、菝契属(SmilaxLinn.)、卫矛属(EuonymusLinn.)、朴属(CeltisLinn)等,这些属的分布范围均延续至温带.旧世界热带有6属,热带亚洲(印度-马来西亚)分布有7属,有山胡椒属(LinderaThunb.)、构属(BroussonetiaL′Herit. Ex Vent.)、鸡矢藤属(PaederiaLinn.)、绞股蓝属(GynostemmaBl.)、清风藤属(SabiaColebr.)、葛藤属(PuerariaDC.).这些热带属分布严格限于热带地区的很少,大多数是由热带扩散到亚热带、甚至温带的分布属,许多热带大属所含的种数很多.
2.3.3 温带分布 是指分布于欧洲、亚洲和北美洲温带地区的属,包括北温带分布、东亚和北美洲间断、东亚(东喜马拉雅-日本)等类型.青檀群落中温带分布属有52个,占总属数的55.3%,所占比例高于中国比例(49.8%)[22],表明该植物区系的温带性质较明显.其中,北温带成分最丰富,北温带分布及其变型共有24属,占总属数的25.5%,如栎属(QuercusLinn.)、榆属(UlmusLinn.)、槭属(AcerL.)、盐肤木属(RhusLinn.)、栒子属(CotoneasterMedik.)等.其它温带分布类型共有28属,占总属数29.8%.
2.3.4 中国特有 是指分布区限于中国境内,或者主要分布于中国境内而稍微超出国界,它能反映一个地区植物区系的基本特征.大贵寺森林公园青檀群落含特有属4个,它们都是单型属或少种属,如水杉属(MetasequoiaMiki ex Hu et W. C. Cheng)、青檀属(PteroceltisMaxim.)是单种属,杉木属(CunninghamiaR.Br.)和牛鼻栓属(FortuneariaRehd. Et Wils.)是少种属.
该青檀群落包含了除温带亚洲、中亚分布外的其它中国种子植物属分布类型,说明其地理成分复杂、多样化.统计结果表明,热带分布属植物所占比例为40.4%;温带分布属植物所占比例为55.3%,中国特有属植物所占为4.3%(表3).可见该青檀群落以温带分布属占一定优势,其次为热带分布属,不过许多热带分布属的分布范围延伸至温带,说明该青檀群落种子植物属的区系成分表现出温带性质的特点.
2.4 物种多样性分析
物种多样性是群落的种类、个体总数及均匀度的综合概念,反映了群落功能及组织特征[23].青檀群落乔木层、灌木层和草本层的物种丰富度指数、Simpson指数、Shannon-Wiener指数和Pielous均匀度指数(表4).
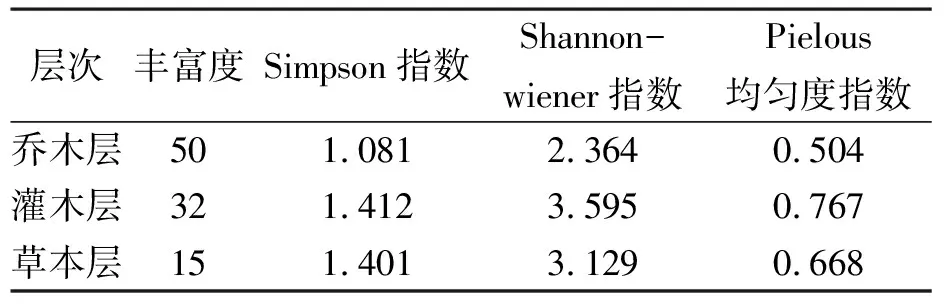
表4 青檀群落三个层次的物种多样性水平
乔木层的丰富度指数最大,即物种数最多,草本层物种数最少.丰富度指数与取样面积关系密切,在进行多样性水平判断时常作为参考.除丰富度指数外,其它多样性指数的变化规律一致,Simpson指数、Shannon-Wiener指数、Pielous均匀度指数均是灌木层>草本层>乔木层.Shannon-Wiener指数是表示群落中物种丰富程度的重要指标,应用较为普遍[24],Pielous均匀度指数反映群落均匀度,可表明群落中物种定量指标的差异程度[25].以Shannon-Wiener指数作为评价的主要指标,结合Pielous均匀度指数,可知三个层次中,灌木层物种多样性水平最高,物种分布均匀,乔木层虽然物种丰富度指数最大,但优势种明显,物种分布不均匀,因而多样性水平较低.
3 讨 论
种子植物属区系分析结果表明,青檀群落以温带分布类型为主,其次为热带分布类型,这与大贵寺国家森林公园地处北亚热带的地理位置是相符的.覃文更[13]所研究的广西木论自然保护区的青檀群落区系成分以热带分布型为主,说明青檀具有很强的适应性,能适应不同气候条件形成不同区系成分的群落类型,反映地理环境条件特点.
青檀群落垂直结构具有层次性,可分为乔木层、灌木层、草本层、层间植物等结构,乔木层和灌木层中青檀占明显优势,说明其对群落的形成及其特征具有重要作用.青檀群落三个层次中,灌木层物种多样性水平最高,其次为草本层,乔木层物种多样性水平最低,与其它地区青檀群落不同层次多样性的规律一致[13].究其原因,与乔木层优势种明显有一定关系,虽然在调查样地中,乔木层物种种数最多,但均匀度指数低,多数种类为偶见种,因而物种多样性指数小;乔木层中的青檀树冠结构不致密,疏荫的环境对灌木层物种以及乔木幼苗幼树的生长有利,因而灌木层物种多样性水平较高.
不过与广西木论自然保护区青檀群落三个层次中多样性指数(Shannon-Wiener指数)[13]相比,大贵寺国家森林公园青檀群落三个层次的物种多样性指数更大,这可能与大贵寺国家森林公园青檀群落发展时间长、受到较好的保护有关,大贵寺国家森林公园游人量不大,且群落取样地点距离游人集中的游览点较远;另一方面可能与取样面积略大有关;再者,可能大贵寺青檀群落的生境条件更为多样化,Shannon-Wiener指数对群落生境的差异敏感[24],因而反映群落物种多样性更高.
青檀是我国特有的、具有较高经济价值的珍贵植物资源,目前国内成片分布的青檀野生群落已不多见,在大贵寺国家森林公园仍有较大面积的集中分布,形成青檀优势种群落,实为难得,应引起重视和加强保护.
第一,加强青檀保护生物学的研究,加强青檀的就地抚育.野外调查发现,虽然青檀大树具有一定的结实量,但幼苗数量并不丰富,说明青檀种子在野外萌发率不高,这可能与种子的生理性休眠
有关,须低温层积处理才能打破休眠[2],因此可加强野外就地抚育,提高种子萌发率,促进幼苗生长,从而促进种群发展.第二,进一步开展对青檀群落的的演替机制的研究,加强群落保护,维持群落的稳定性和多样性.第三,加强森林公园管理以及青檀群落生境的保护,划定青檀保护区.第四,坚持开展植物保护和管理知识的教育,使全社会支持和参与保护工作.
参考文献:
[1] 中国科学院中国植物志编辑委员会.中国植物志:第22卷[M].北京:科学出版社,1998.
[2] 洑香香,方升佐,杜艳.青檀种子休眠机理及发芽条件的探讨[J].植物资源与环境学报, 2002,11(1): 9-13.
[3] 方升佐,朱梅,唐罗忠,等.不同种源青檀种子的营养成分及种子活力的差异[J].植物资源与环境学报,1998,7(2):16-22.
[4] Fang S Z, Song L Y, Fu X X. Effects of NaCl stress on seed germination, leaf gas exchange and seedling growth ofPteroceltistatarinowii[J].Journal of Forestry Research, 2006,17(3):185-188.
[5] 洑香香,方升佐,汪红卫,等.青檀一年生播种苗的年生长规律[J].南京林业大学学报:自然科学版,2001,25(6):11-14.
[6] 张兴旺,张小平,杨开军,等.珍稀植物青檀叶解剖结构及其生态适应性特征[J].植物研究,2007,27(1):38-42.
[7] 李光友,方升佐,吕家驹,等.立地条件对青檀人工林生物生产力及檀皮产量的影响[J].南京林业大学学报:自然科学版,2001,25(4):49-53.
[8] 方升佐,李光友,洑香香.立地条件对青檀檀皮中矿质元素含量的影响[J].林业科学,2002,38(1):8-14.
[9] 方升佐,李光友,李同顺,等.经营措施对青檀人工林生物量及檀皮产量的影响[J].植物资源与环境学报,2001,10(1):21-24.
[10] 方升佐,崔同林,虞木奎.成土母岩和条龄对青檀檀皮质量的影响[J].北京林业大学学报,2007,29(2):122-127.
[11] Fang S Z,Li G Y,Fu X X.Biomass production and bark yield in the plantations of Pteroceltis tatarinowii[J]. Biomass & Bioenergy, 2004,26(4): 319-328.
[12] 王文静,何雅蔷.宝天曼自然保护区青檀林结构特征与物种多样性研究[J].河南农业大学学报,2001,35(1):364-367.
[13] 覃文更,韦国富,谭卫宁.广西木论自然保护区青檀群落特征及其多样性研究[J].广西林业科学,2004,33(3):126-129.
[14] Chai X Y, Chen S L, Xu W. Using inter-simple sequence repeat markers to analyze the genetic structure of naturalPteroceltistatarinowiipopulations and implications for species conservation[J]. Plant Syst Evol, 2010,285:65-73.
[15] 时宝茹,张辉勇.大贵寺国家森林公园青檀资源保护与对策[J].湖北林业科技,2006,(4):54-56.
[16] 李博.生态学[M].北京:高等教育出版社,2000.
[17] 吴征镒.中国种子植物属的分布区类型[J].云南植物研究,1991(增刊):1-139.
[18] 吴征镒.中国种子植物属的分布区类型增补和勘误[J].云南植物研究,1993(增刊):141-178.
[19] 郭泉水,江洪,王兵,等.中国主要森林群落植物生活型谱的数量分类及空间分布格局的研究.生态学报,1999,19(4):573-574.
[20] 雷泞菲,苏智先,宋会兴,等.缙云山常绿阔叶林不同演替阶段植物生活型谱比较研究.应用生态学报, 2002,13(3):267-270
[21] 汪殿蓓,王彩云,暨淑仪,等.野生仙湖苏铁(Cycasfairylakea)群落特征研究[J].北京林业大学学报,2004,26(6):12-18.
[22] 王荷生.中国植物区系的性质和各成分间的关系[J].云南植物研究,2000,22(2):119-126.
[23] 毕润成,成亚丽,尹大泽,等.吕梁山南端白皮松的群落特征及其多样性的研究[J].植物研究,2002,22(3): 366-372.
[24] 彭少麟,陈章和.广东亚热带森林群落物种多样性[J].生态科学,1983,(2):98-104.
[25] 钱迎倩,马克平.生物多样性研究的原理方法[M].北京:中国科学技术出版社,1994.
